大部分神经元的树突并不光滑,来自轴突的输入在扣节(bouton)处与树突棘(dendritic spines)处精准连接形成突触。在大脑皮层的传统切片染色或近年来的电子显微镜三维重建研究中,相对于细胞体、轴突、树突轴等结构,树突棘占据了相当可观的体积,累计可以超过20%的皮层区域[1]。该项研究中75%的突触涉及树突棘。树突棘的概念(图6.1),及其对于学习和记忆的重要性可以追溯到一百多年前,西班牙的首位诺贝尔生理学与医学奖得主、少年期顶住父亲压力研习绘画的圣地亚哥·拉蒙-卡哈尔(Santiago Ramón y Cajal)在神经元的染色图像中首次观察到树突棘[2]。除了兴奋性的锥体神经元,兴奋性的中间神经元棘状星形细胞(spiny stellate cells)、抑制性的中间神经元中反应较慢的Martinotti细胞(表达生长抑素somatostatin,SST/SOM细胞,上一节图5.1),以及小脑的浦肯野细胞(Purkinje cells)也有树突棘。
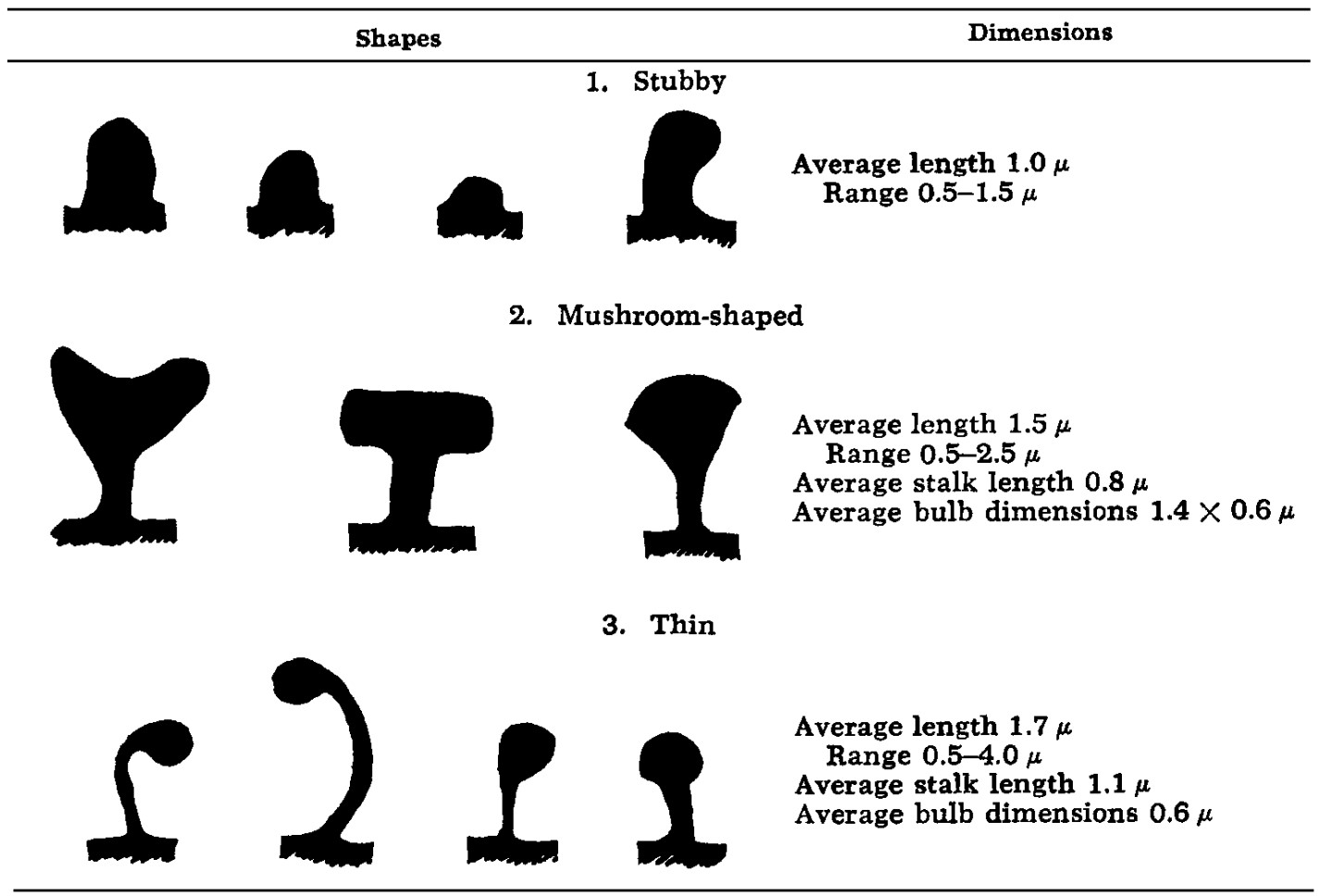
图6.1 大鼠大脑皮层树突棘的形状和长宽。树突棘遍布锥体神经元(本书第4节)和棘状星形细胞的树突。来自[3]。

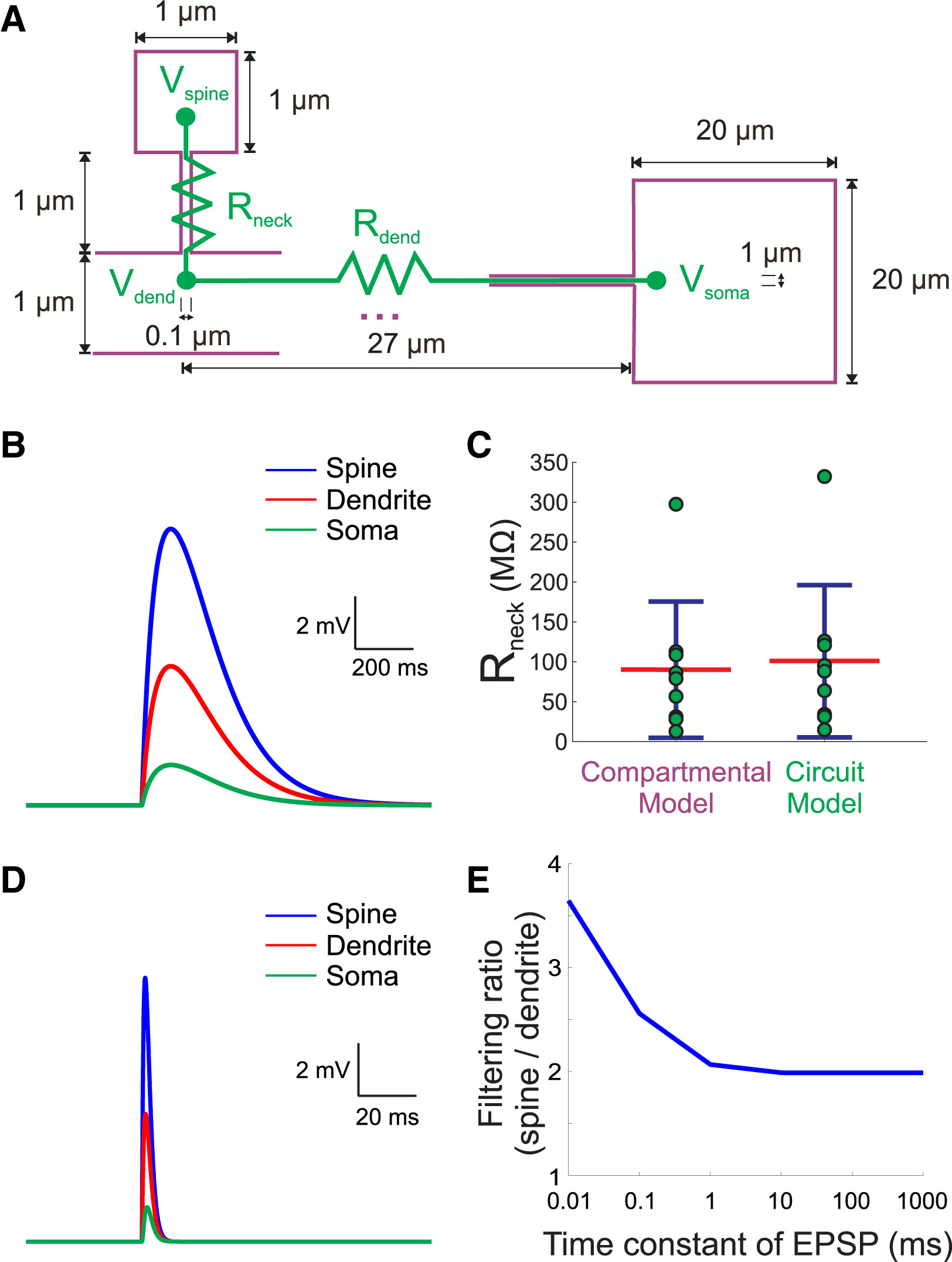
图6.3 对树突棘(Rneck)电阻的估算。(A)一个体外培养的小鼠海马神经元的电阻分别根据电路模型(绿色)和腔体模型(紫色)估算,单个树突棘颈部的电阻中位数都在100兆欧以上(C)。在腔体模型估算中,该海马神经元树突棘头部1微米x1微米的尺寸比图6.1小鼠大脑皮层神经元树突棘的尺寸大;胞体为20微米x20微米。(B)在根据腔体模型估算树突棘电阻时所作的电压模拟,小图比例尺显示y轴2mV对应的高度和x轴200ms对应的宽度。树突轴电阻(Rdend)采用文献报道的平均108兆欧。图示的本次模拟得出树突棘颈部电阻为95兆欧,(C)如图(B),左侧为根据腔体模型,树突轴电阻在63-153 MΩ范围内的多次模拟,得出树突棘颈部电阻范围64-146 MΩ。右侧为根据电路模型得出的63-153 MΩ。(D)在快速兴奋性突触后电位(EPSP)为1 ms时间常数下,像(B)一样模拟腔体模型,获得树突棘对电信号的过滤比。图示的1ms时间常数模拟已经与更慢的EPSP结果接近(D)。可以看到从树突棘、树突轴到细胞体,电压显著减小。(E)如图(D)的不同时间常数多次模拟(τ=0.01, 0.1, 1, 10, 100, 1000 ms),得出树突棘相对于树突轴对膜电压的过滤比。来自[5]。
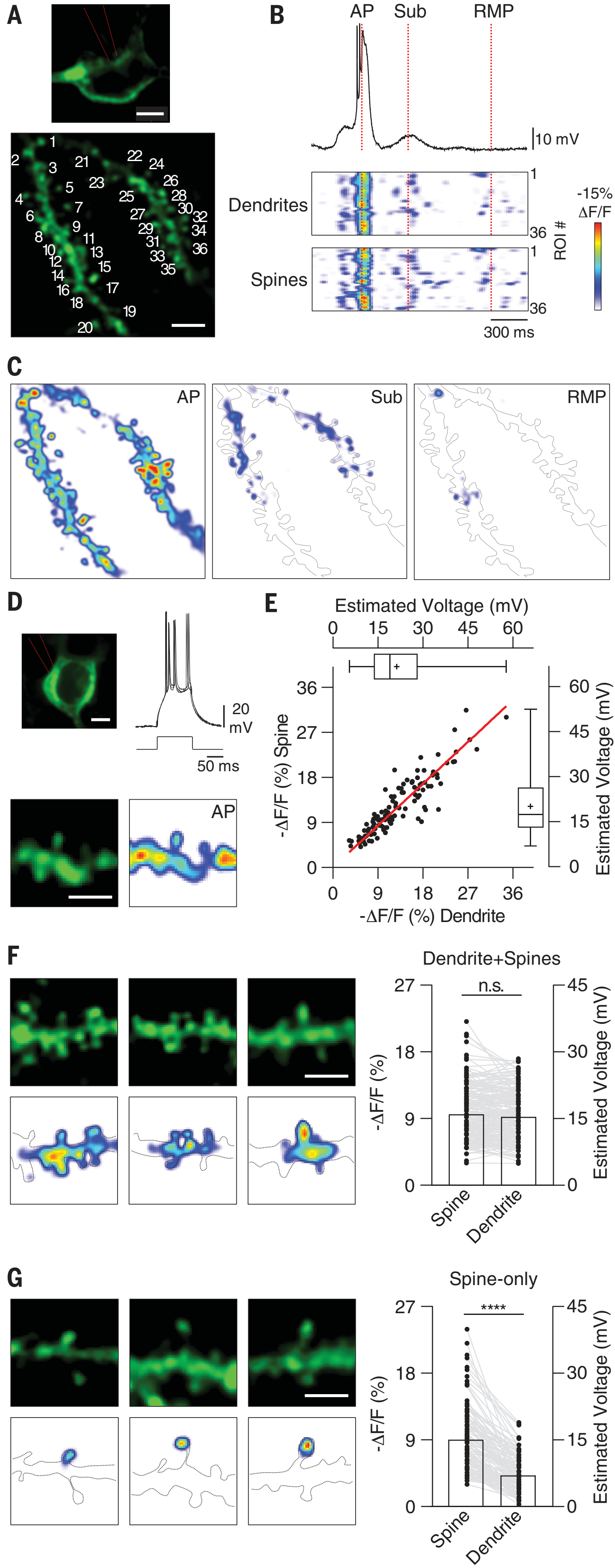
图6.4 基因改造的小鼠神经元表达对电压敏感的荧光蛋白,可以直接在树突棘和树突轴中显示数十毫伏的电压变化。AP,train of action potentials,动作电位系列;Sub,subthreshold dipolarization,未达到动作电位阈值的去极化。RMP,resting membrane potential,静息膜电位。子图(D)的动作电位系列由三次100毫秒的脉冲(300pA电流)激发。来自[6]。
如图6.4 C、F、G子图所示,实验小鼠在光遗传学刺激下,神经元可以只在树突脊内或树突轴特定区段内被激活,而不是整根树突轴激活[6]。动作电位时,神经元细胞整体进行数据更新(图6.4C、D)。
上一节提到的中间神经元,大篮状细胞经常在细胞核周边的胞体上有较大的轴突对锥体神经元施加抑制信号,而枝状细胞通常在轴突起始位置施加抑制信号。
虽有大量树突棘的过滤,神经元细胞核周边的电压仍然有毫伏级。笔者在后面章节推测,树突棘的电活动可能在非侵入性的脑电图中(通常在1-100微伏)已经直接被检测到了。
参考文献:
[1] Braitenberg, Valentino, and Almut Schüz. 1998. Cortex: Statistics and Geometry of Neuronal Connectivity. Cortex: Statistics and Geometry of Neuronal Connectivity. Berlin, Heidelberg: Springer Berlin Heidelberg. https://doi.org/10.1007/978-3-662-03733-1.
[2] Ehrlich, B. The Brain in Search of Itself : Santiago Ramón y Cajal and the Story of the Neuron. (Farrar, Straus and Giroux, 2022).
[3] Peters, A. & Kaiserman-Abramof, I. R. The small pyramidal neuron of the rat cerebral cortex. The perikaryon, dendrites and spines. Am. J. Anat. 127, 321–55 (1970).
[4] Schüz, A. Prenatal development and postnatal changes in the guinea pig cortex: microscopic evaluation of a natural deprivation experiment. II. Postnatal changes. J. Hirnforsch. 22, 113–27 (1981).
[5] Kwon, T., Sakamoto, M., Peterka, D. S. & Yuste, R. Attenuation of synaptic potentials in dendritic spines. Cell Rep. 20, 1100–1110 (2017).
[6] Cornejo, V. H., Ofer, N. & Yuste, R. Voltage compartmentalization in dendritic spines in vivo. Science (80-.). (2021) doi:10.1126/science.abg0501.

0
推荐
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}