面对大量信息的轰炸,大脑每天都要应对的一个关键问题是,要存储哪些,存在哪里。关于大脑的记忆,并没有形而上学的玄妙,而是可归结到神经元细胞和它们的连接模式,其物理基础最终还是取决于单个突触。
一个流行了很久的祖母悖论,基于传统的神经细胞一层层自上而下理念,是说如果失去了所谓的单一祖母细胞,会让你忘记自己的祖母。事实上,伴随着丰富的体验,许多与不同感官相连的神经元都可能与一个人的祖母有关,这种各有侧重的多重备份下,我们不必担心失去一个无法更新的祖母细胞。祖母悖论的流行,也反映了神经元层级观念的根深蒂固,而忽略了记忆在同层和不同层神经元、不同神经模块之间相互的不完全备份。
从记忆的物理过程来说,新的经历要么被写在以前已编码相关经历的神经元上,要么在这些神经元附近招募更多的神经元进行存储。任何一个当时比较活跃的神经元都可以参与记忆,形成一个多感官的时空片段。这种持续竞争的过程,天然地导致更久远的事件在检索时只激活少量神经元及其突触。而一些事件在一开始就占据了更多的神经元细胞(图14.1),且倾向于被回放,比如激发了情绪的事件 。记忆的整理过程,我们在睡眠的章节还会继续讲。
不仅是事件,规律也在单细胞水平(可能在单个树突棘,本书后面章节内容)存储与更新。即使所有相关的连接都消失了,只要神经元和蛋白质还在那里,记忆痕迹也可以被直接激活(激光照射小鼠模型中光敏蛋白标记的神经元)[1,2]。从这个意义上说,我们可能确实记得我们曾经经历的一切; 而记忆的读取则更多的是一个检索问题,包括把尘封的记忆从更快更强的信号覆盖下释放出来,然后还涉及一些补全。
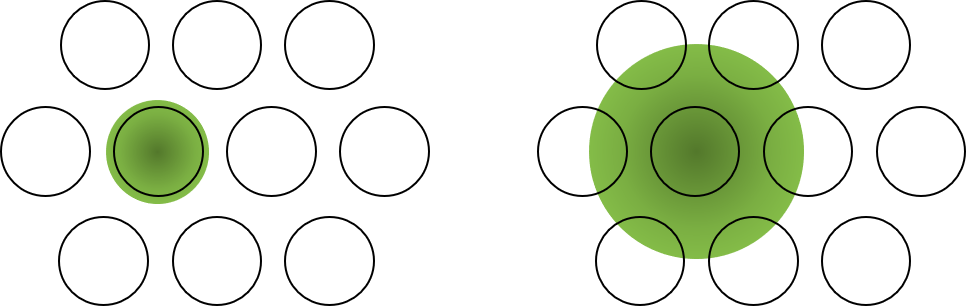
图14.1 一个信号的扩散范围或波可能决定了分配给每个事件的神经元的数量。例如,一个非常情绪化的事件可能会招募太多的神经元。而一些精神类药物可能通过改变扩散时间影响了涉及的神经元数量。来源:作者自绘
前面章节讲到,除了经典的毫秒级的点对点突触间化学信号变电信号的传输外,还存在较快的缝隙连接(以纳秒为单位,比如PV中间神经元的协调) ,以及更慢一些的扩散分子或囊泡,即当蛋白质泄漏到目标突触时发生的容积传递(volume transmission),例如多巴胺被释放到神经元之外。
前文讲到的祖母悖论,笔者认为使记忆更强的一个简单办法就是占用更多的细胞。当某个情绪性过强的记忆占据了过多细胞后,会导致其被过多的激活,这会导致诸如PTSD的症状。发育生物学中常见的图灵斑图(图14.2)[3],包括豹纹、指纹的形成,通常出现在反应扩散系统中,是由局部激活和长程抑制一起形成的圆圈或条纹。脑内可能还存在更多源头的复杂干涉。后面章节提到的网格细胞的发育可能也涉及图灵班图。除了圈地(扩散到更多神经元),笔者猜测图灵班图式的扩散波或电信号可能也是一种产生时序规律的方式,时空得到后面章节才讲。
抗抑郁的monoamine oxidase inhibitors(MAOi)减慢人脑对去甲肾上腺素、血清素、或多巴胺的代谢,使得递质的存在时间更长,可能让激活圈更大(图14.1),从而加强了当前事件的影响。
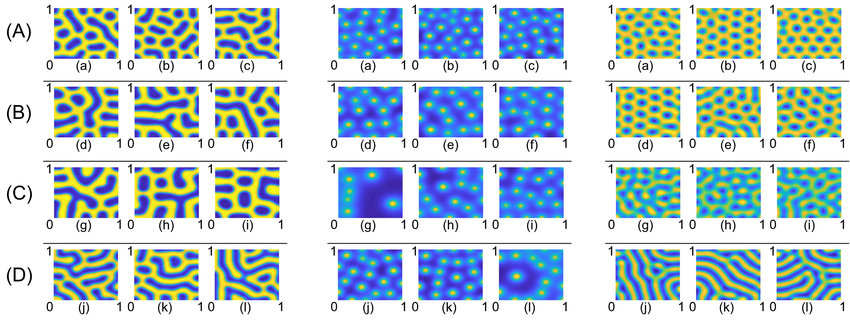
图14.2 图灵班图的形成过程。反应源及其远端抑制的不同参数可以形成不同的斑点或条纹。来自[2]
针对单一事件或规则,记忆分配的神经元并非单一,不同事件对应的记忆神经元也存在交集。例如两个时间上相近的事件,其分配的用于存储的神经元集合,即记忆印痕(memory engram),存在重叠[4]。小鼠被训练着先根据一种规律,再采用另一种规律对视觉刺激作出反应,也会发现用于记忆两种规则的神经元集合存在部分重叠[5]。记忆在细胞层面的神经元复用,既高效、节俭,又天然地有逻辑框架。大脑的记忆可以比作超市的货架,有大的分区,也有相邻物品之间的联系。
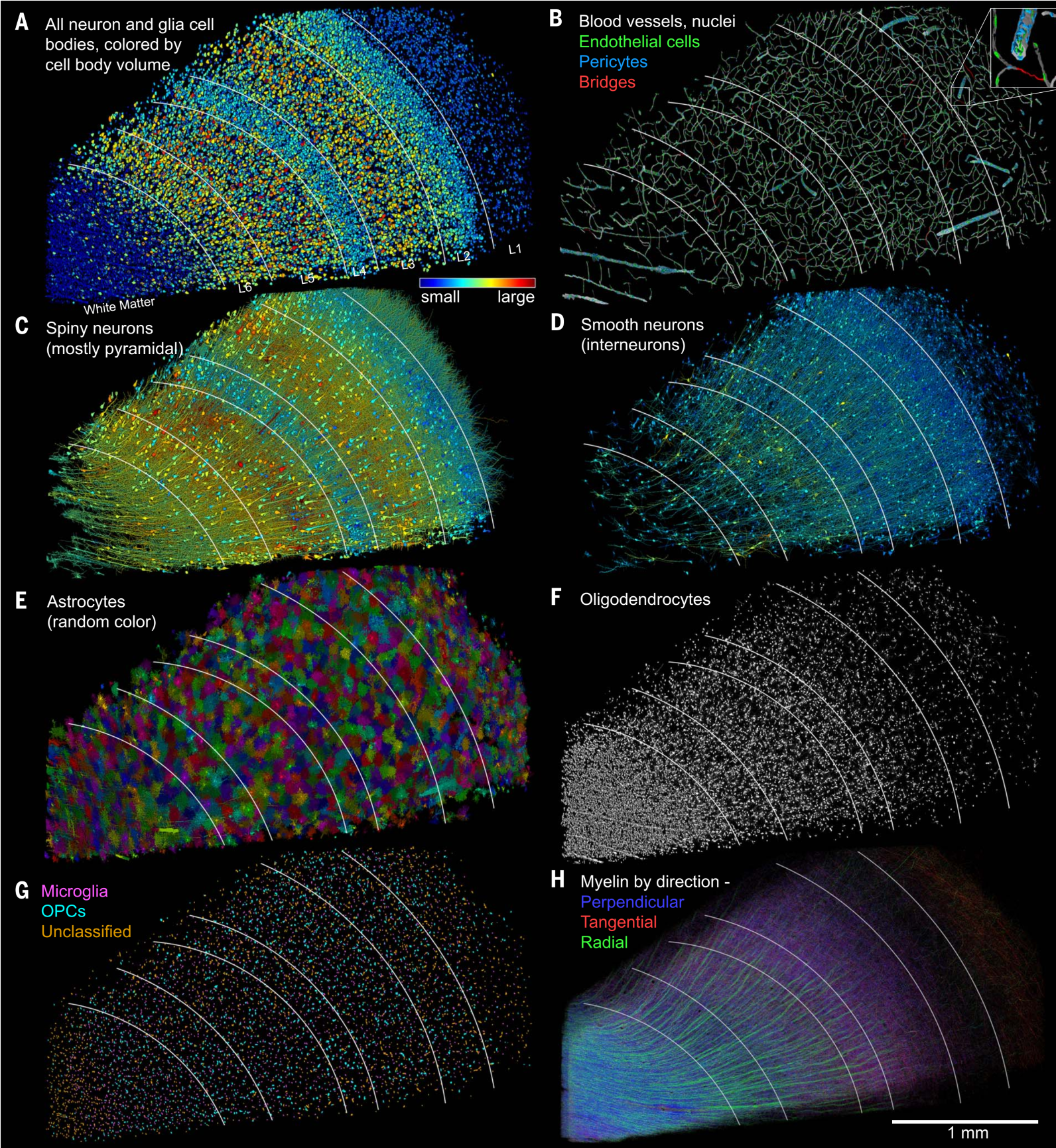
图14.3 对人脑颞叶皮层1立方毫米样本的电子显微镜三维重建结果。(A)样本中所有49080个神经细胞和胶质细胞。根据细胞体(细胞核所在位置,不包括树突、轴突延伸)由小到大显示为蓝色至红色。白线分隔的L1至L6即第一层至第六层。(B)脑血管和相关8136个细胞。(C)10531个含树突脊、预计大部分为兴奋性的锥体神经元的细胞,也按细胞体大小显示为不同颜色。(D)4688个基本没有树突脊、预计为抑制性的中间神经元的细胞。(E)5474个星形胶质细胞。(F)20139个可以包裹轴突、形成髓鞘(myelin,子图H)的少突胶质细胞(oligodendrocytes),可能还有更多。(G)6702个小胶质细胞及细胞。(H)髓鞘化的轴突。大脑皮层白质中大部分髓鞘与皮层垂直分布。来自[6],本书第4节已部分呈现。
除了神经元,遍布大脑皮层的星形胶质细胞(astrocytes)(图14.3)也参与记忆印记的形成,这也是后文阿尔茨海默症会提到的一大细胞类型。(恐惧记忆)学习诱导了海马区的星形胶质细胞表达cFos蛋白,而这些辅助性细胞与印痕神经元相作用,再次激活这些星形胶质细胞也可以再现恐惧反应[7,8]。
一项最新的研究显示,星形胶质细胞在神经元突触周围形成包裹,这种由星形胶质细胞形成的小空间对钙离子浓度的保持、神经元之间信号整合都有作用[9]。
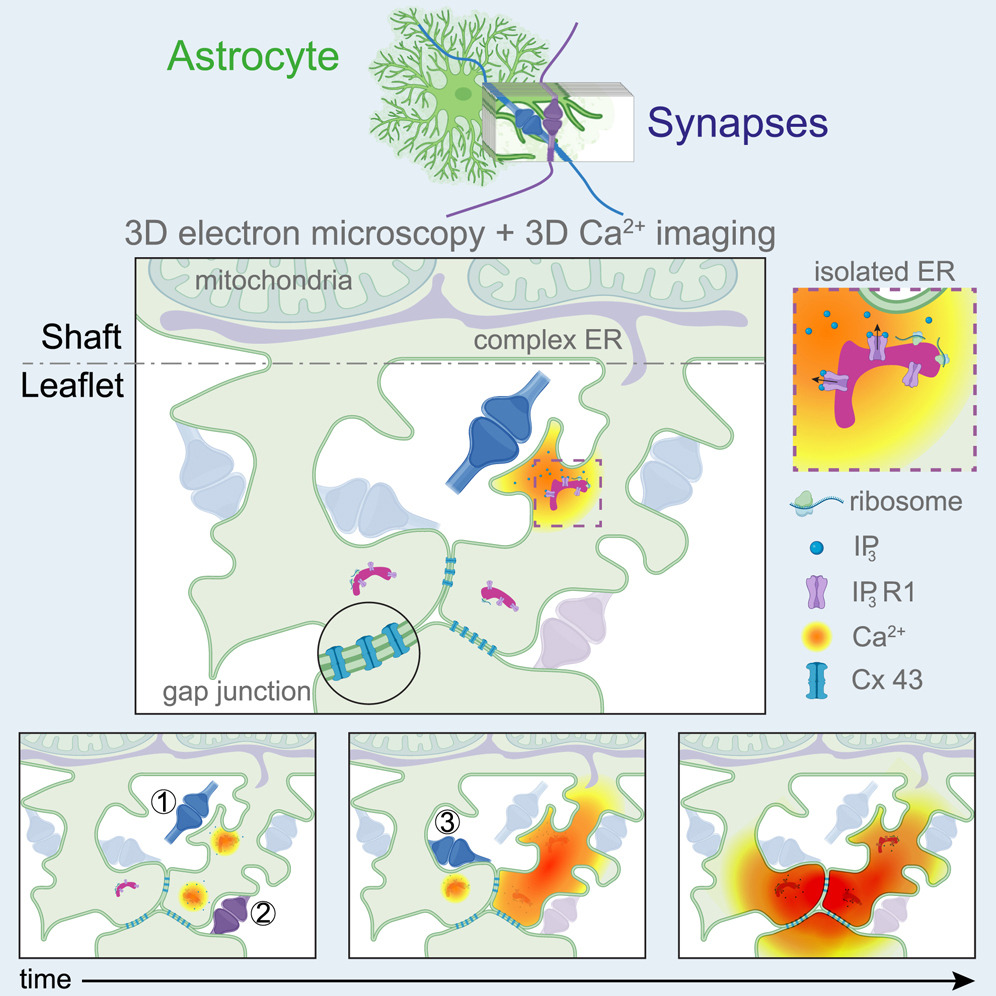
图14.4 星形胶质细胞分支末梢形成小叶结构,包裹在一个或多个突触周围。星形胶质细胞的小叶之间还会建立缝隙连接。小叶包裹区域的钙离子浓度可以被局部改变。来自[9]。
老年小鼠在少突胶质细胞(oligodendrocytes)对神经元的髓鞘化(图14.3F,H)方面也显示了衰退[10]。表达小清蛋白(快速结合钙离子)的抑制性中间神经元的轴突是大脑皮层和海马中被髓鞘化的较大一部分[11,12]。老年小鼠中髓鞘化的衰退预计影响了电信号的保真,且因为涉及这些中间神经元,可能也使得记忆更为碎片化,难以在同层的较大尺度上形成新的组合。
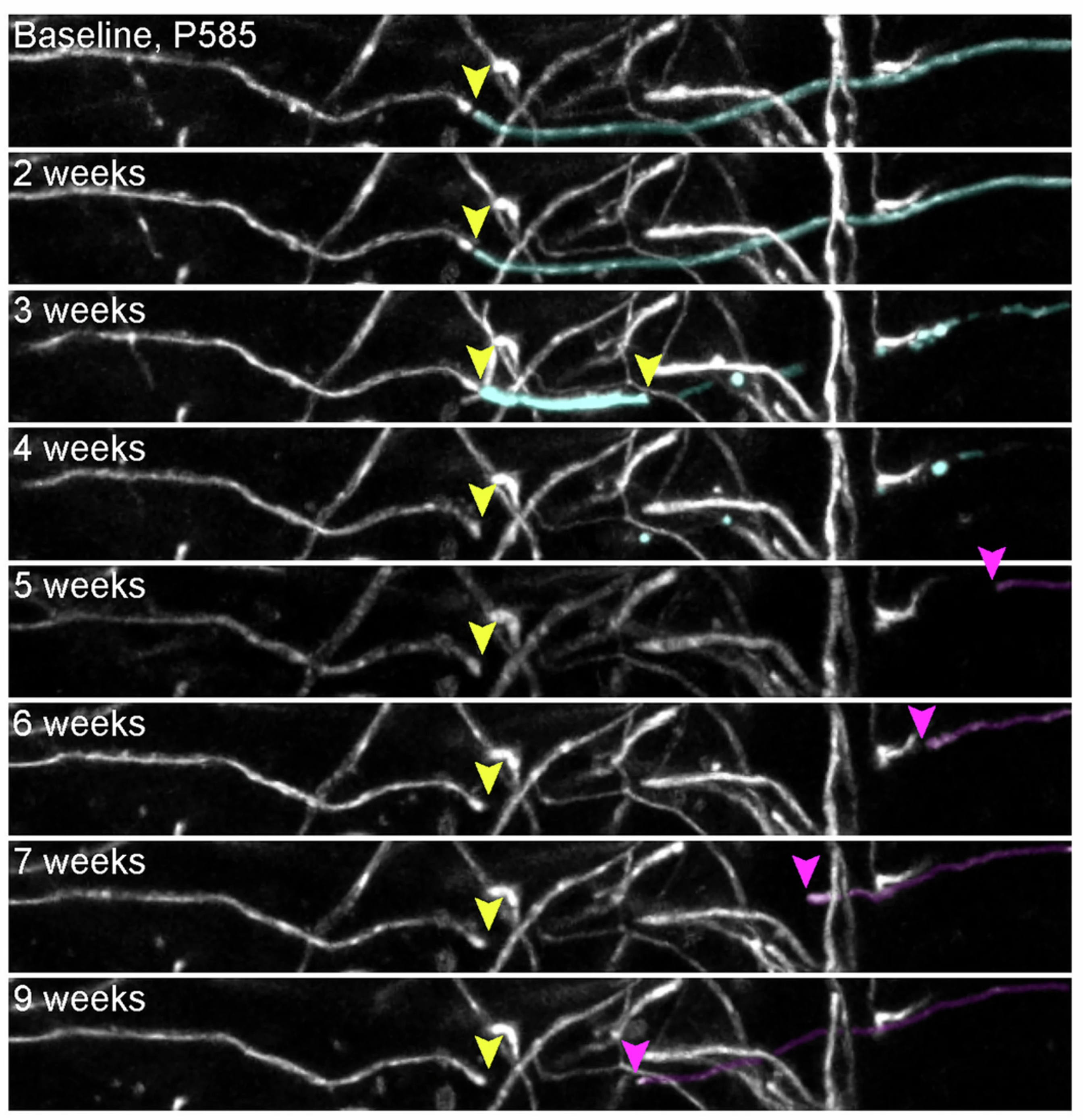
图14.5 一例髓鞘化丢失(黄色)和部分修复(紫色)的例子。来自[10]。
确立了记忆在神经细胞水平、而不是更泛化的所谓网络的存储,后面章节才能更深入的展开。有更新阈值和索引阵列的准确存储,是准确建图与复用,进而开展因果推断的基石。
参考文献:
[1] Josselyn, Sheena A., and Susumu Tonegawa. (2020). “Memory Engrams: Recalling the Past and Imagining the Future.” Science (New York, N.Y.) 367 (6473): eaaw4326.
[2] Ryan, Tomás J., and Paul W. Frankland. (2022). “Forgetting as a Form of Adaptive Engram Cell Plasticity.” Nature Reviews Neuroscience 23 (3): 173–186.
[3] Alexey Kazarnikov, & Heikki Haario. (2020). Statistical approach for parameter identification by Turing patterns. Journal of Theoretical Biology, 501, 110319–110319.
[4] Josselyn, Sheena A., and Susumu Tonegawa. (2020). “Memory Engrams: Recalling the Past and Imagining the Future.” Science (New York, N.Y.) 367 (6473): eaaw4326.
[5] Reinert, Sandra, Mark Hübener, Tobias Bonhoeffer, and Pieter M. Goltstein. (2021). “Mouse Prefrontal Cortex Represents Learned Rules for Categorization.” Nature 593 (7859): 411–417.
[6] Shapson-Coe, A., Januszewski, M., Berger, D. R., Pope, A., Wu, Y., Blakely, T., Schalek, R. L., Li, P. H., Wang, S., Maitin-Shepard, J., Neha Karlupia, Sven Dorkenwald, Sjostedt, E., Leavitt, L., Lee, D., Troidl, J., Collman, F., Bailey, L., Fitzmaurice, A., & Kar, R. (2024). A petavoxel fragment of human cerebral cortex reconstructed at nanoscale resolution. Science, 384(6696). https://doi.org/10.1126/science.adk4858
[7] Williamson, M. R., Kwon, W., Woo, J., Ko, Y., Maleki, E., Yu, K., Murali, S., Sardar, D., & Deneen, B. (2024). Learning-associated astrocyte ensembles regulate memory recall. Nature. https://doi.org/10.1038/s41586-024-08170-w
[8] Dewa KI, Kaseda K, Kuwahara A, Kubotera H, Yamasaki A, Awata N, Komori A, Holtz MA, Kasai A, Skibbe H, Takata N, Yokoyama T, Tsuda M, Numata G, Nakamura S, Takimoto E, Sakamoto M, Ito M, Masuda T, Nagai J. (2025). The astrocytic ensemble acts as a multiday trace to stabilize memory. Nature. 648(8092):146-156.
[9] Benoit L, Hristovska I, Liaudet N, Jouneau PH, Fertin A, de Ceglia R, Litvin DG, Di Castro MA, Jevtic M, Zalachoras I, Kikuchi T, Telley L, Bergami M, Usson Y, Hisatsune C, Mikoshiba K, Pernet-Gallay K, Volterra A. (2025) Astrocytes functionally integrate multiple synapses via specialized leaflet domains. Cell. 188(23):6453-6472.e16.
[10] Call CL, Neely SA, Early JJ, James OG, Zoupi L, Williams AC, Xu YKT, Chandran S, Lyons DA, Monk KR, Bergles DE. (2026) Flexible ensheathment of axons enables myelination of complex CNS networks. Nature doi.org/10.1038/s41586-026-10312-1
[11] Micheva KD, Wolman D, Mensh BD, Pax E, Buchanan J, Smith SJ, Bock DD. A large fraction of neocortical myelin ensheathes axons of local inhibitory neurons. Elife. 2016 Jul 6;5:e15784.
[12] Stedehouder J, Couey JJ, Brizee D, Hosseini B, Slotman JA, Dirven CMF, Shpak G, Houtsmuller AB, Kushner SA. Fast-spiking parvalbumin interneurons are frequently myelinated in the cerebral cortex of mice and humans. Cereb Cortex. (2017) 27(10):5001-5013.

0
推荐
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}