[1] Ardila, A., Bernal, B. & Rosselli, M. How localized are language brain areas? a review of brodmann areas involvement in oral language. Arch. Clin. Neuropsychol. 31, 112–122 (2016). [2] K, Thompson, Gibbings A, Shaw J, Ray L, Hébert G, De Koninck J, and Fogel S. 2021. “Sleep and Second-Language Acquisition Revisited: The Role of Sleep Spindles and Rapid Eye Movements.” Nature and Science of Sleep 13 (October): 1887–1902. https://doi.org/10.2147/NSS.S326151.
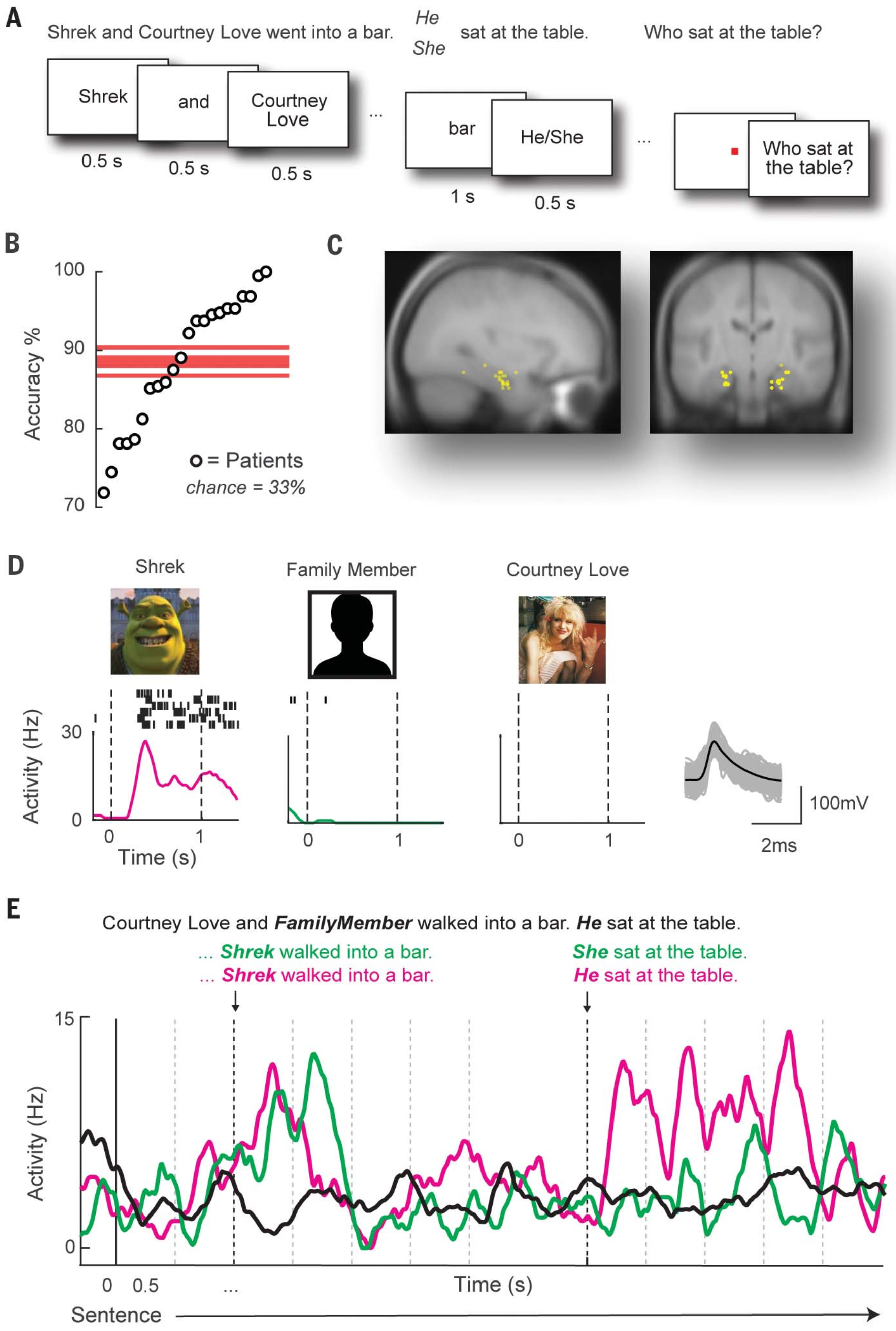
[3] Dijksterhuis, D. E., Self, M. W., Possel, J. K., Peters, J. C., van Straaten, E. C. W., Idema, S., Baaijen, J. C., van der Salm, S. M. A., Aarnoutse, E. J., van Klink, N. C. E., van Eijsden, P., Hanslmayr, S., Chelvarajah, R., Roux, F., Kolibius, L. D., Sawlani, V., Rollings, D. T., Dehaene, S., & Roelfsema, P. R. (2024). Pronouns reactivate conceptual representations in human hippocampal neurons. Science, 385(6716), 1478–1484.
[4] Johnson, Elliott Gray, Lindsey Mooney, Katharine Graf Estes, Christine Wu Nordahl, and Simona Ghetti. 2021. “Activation for Newly Learned Words in Left Medial-Temporal Lobe during Toddlers’ Sleep Is Associated with Memory for Words.” Current Biology 31 (24): 5429-5438.e5. https://doi.org/10.1016/j.cub.2021.09.058.
[5] Boros, Marianna, Lilla Magyari, Dávid Török, Anett Bozsik, Andrea Deme, and Attila Andics. 2021. “Neural Processes Underlying Statistical Learning for Speech Segmentation in Dogs.” Current Biology 31 (24): 5512-5521.e5. https://doi.org/10.1016/j.cub.2021.10.017.
[6] Tang, J.C.Y., Paixao, V., Carvalho, F. et al. Dynamic behaviour restructuring mediates dopamine-dependent credit assignment. Nature626, 583–592 (2024). https://doi.org/10.1038/s41586-023-06941-5
[7] Blum, F., Paschen, L., Forkel, R. et al. Consonant lengthening marks the beginning of words across a diverse sample of languages. Nat Hum Behav8, 2127–2138 (2024). https://doi.org/10.1038/s41562-024-01988-4
[8] Ullman, M T, and Michael T. Ullman. 2001. “A Neurocognitive Perspective on Language: The Declarative/Procedural Model.” Nature Reviews. Neuroscience 2 (10): 717–26. https://doi.org/10.1038/35094573.
[9] Wei, Xin Paul, Matthew Collie, Bowen Dempsey, Gilles Fortin, and Kevin Yackle. 2022. “A Novel Reticular Node in the Brainstem Synchronizes Neonatal Mouse Crying with Breathing.” Neuron 110 (4): 644-657.e6. https://doi.org/10.1016/j.neuron.2021.12.014.
[10] Ghazanfar, Asif A, and Daniel Y Takahashi. 2014. “The Evolution of Speech: Vision, Rhythm, Cooperation.” Trends in Cognitive Sciences 18 (10): 543–53. https://doi.org/10.1016/j.tics.2014.06.004.
[11] Guilleminot, Pierre, and Tobias Reichenbach. 2022. “Enhancement of Speech-in-Noise Comprehension through Vibrotactile Stimulation at the Syllabic Rate.” Proceedings of the National Academy of Sciences 119 (13): e2117000119. https://doi.org/10.1073/pnas.2117000119.
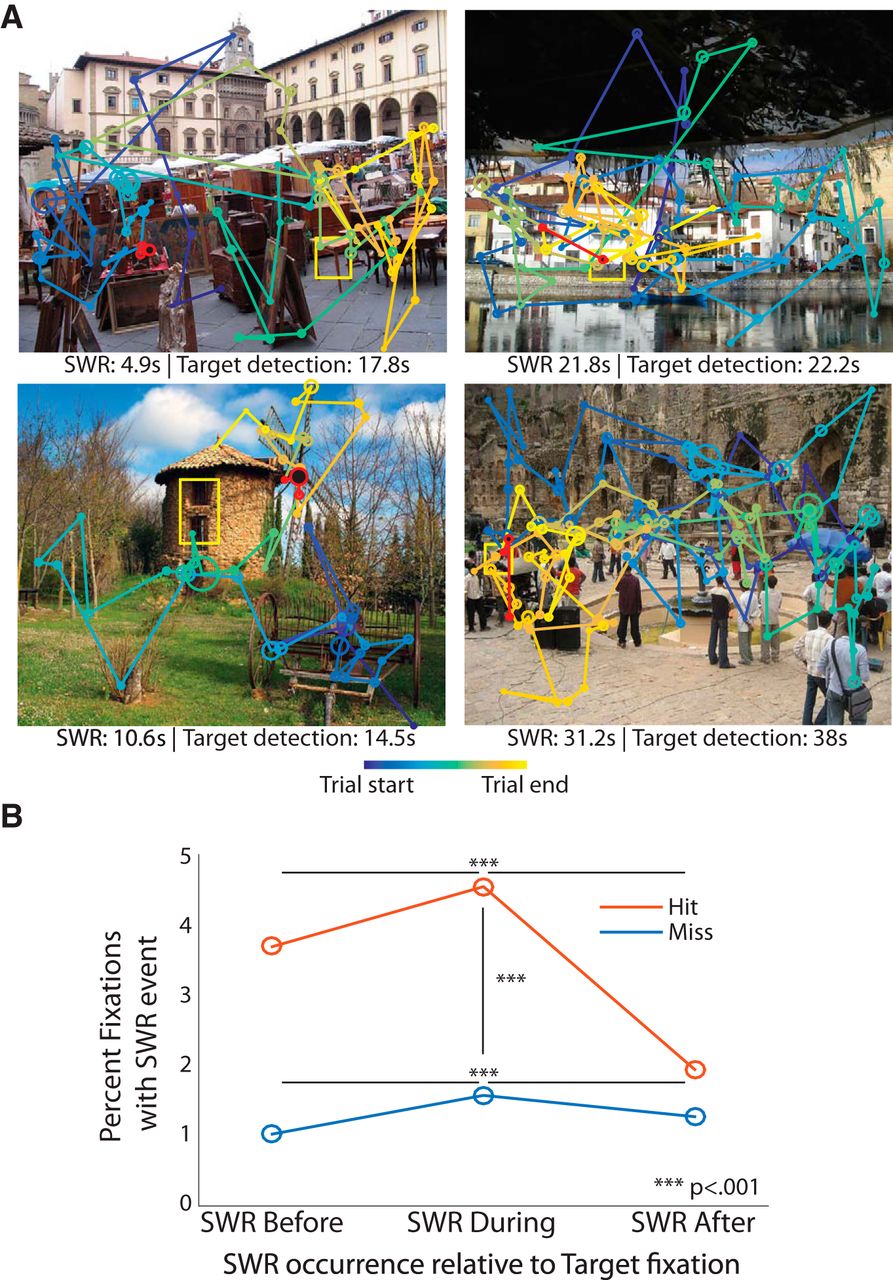
[12] Norman, Yitzhak, Erin M. Yeagle, Simon Khuvis, Michal Harel, Ashesh D. Mehta, and Rafael Malach. 2019. “Hippocampal Sharp-Wave Ripples Linked to Visual Episodic Recollection in Humans.” Science (New York, N.Y.) 365 (6454): eaax1030. https://doi.org/10.1126/SCIENCE.AAX1030.
[13] Castellucci, Gregg A., Christopher K. Kovach, Matthew A. Howard, Jeremy D. W. Greenlee, and Michael A. Long. 2022. “A Speech Planning Network for Interactive Language Use.” Nature 602 (7895): 117–22. https://doi.org/10.1038/s41586-021-04270-z.
[14] Jasmin, Kyle, César F. Lima, and Sophie K. Scott. 2019. “Understanding Rostral–Caudal Auditory Cortex Contributions to Auditory Perception.” Nature Reviews Neuroscience 20 (7): 425–34. https://doi.org/10.1038/s41583-019-0160-2.
[15] Leonard, T. K., J. M. Mikkila, E. N. Eskandar, J. L. Gerrard, D. Kaping, S. R. Patel, T. Womelsdorf, and K. L. Hoffman. 2015. “Sharp Wave Ripples during Visual Exploration in the Primate Hippocampus.” Journal of Neuroscience 35 (44): 14771–82. https://doi.org/10.1523/JNEUROSCI.0864-15.2015.
[16] Killian, Nathaniel J., Michael J. Jutras, and Elizabeth A. Buffalo. 2012. “A Map of Visual Space in the Primate Entorhinal Cortex.” Nature 491 (7426): 761–64. https://doi.org/10.1038/nature11587.
[17] MLR, Meister, and Buffalo EA. 2018. “Neurons in Primate Entorhinal Cortex Represent Gaze Position in Multiple Spatial Reference Frames.” The Journal of Neuroscience : The Official Journal of the Society for Neuroscience 38 (10): 2430–41. https://doi.org/10.1523/JNEUROSCI.2432-17.2018.
[19] Tsu, Jing. 2022. Kingdom of Characters : The Language Revolution That Made China Modern. Riverhead Books.
[20] Kriegeskorte, N. & Wei, X.-X. Neural tuning and representational geometry. Nat. Rev. Neurosci. 22, 703–718 (2021).
[21] Silverstein, Steven M, and Richard Rosen. 2015. “Schizophrenia and the Eye.” Schizophrenia Research. Cognition 2 (2): 46–55. https://doi.org/10.1016/j.scog.2015.03.004.
[22] Trampush, J W, M L Z Yang, J Yu, E Knowles, G Davies, D C Liewald, J M Starr, et al. 2017. “GWAS Meta-Analysis Reveals Novel Loci and Genetic Correlates for General Cognitive Function: A Report from the COGENT Consortium.” Molecular Psychiatry 22 (3): 336–45. https://doi.org/10.1038/mp.2016.244.
[23] Hellard, Stéphanie Le, Yunpeng Wang, Aree Witoelar, Verena Zuber, Francesco Bettella, Kenneth Hugdahl, Thomas Espeseth, et al. 2017. “Identification of Gene Loci That Overlap Between Schizophrenia and Educational Attainment.” Schizophrenia Bulletin 43 (3): 654–64. https://doi.org/10.1093/schbul/sbw085.
[24] Scholl, Benjamin, Clara Tepohl, Melissa A. Ryan, Connon I. Thomas, Naomi Kamasawa, and David Fitzpatrick. 2022. “A Binocular Synaptic Network Supports Interocular Response Alignment in Visual Cortical Neurons.” Neuron 110 (9): 1573-1584.e4. https://doi.org/10.1016/j.neuron.2022.01.023.
[25] Mesulam, M. 1998. “From Sensation to Cognition.” Brain 121 (6): 1013–52. https://doi.org/10.1093/brain/121.6.1013.
Shepherd, Gordon M.G., and Naoki Yamawaki. 2021.
[26]“Untangling the Cortico-Thalamo-Cortical Loop: Cellular Pieces of a Knotty Circuit Puzzle.” Nature Reviews Neuroscience 22 (7): 389–406. https://doi.org/10.1038/s41583-021-00459-3.
[27] Fraenz, Christoph, Caroline Schlüter, Patrick Friedrich, Rex E. Jung, Onur Güntürkün, and Erhan Genç. 2021. “Interindividual Differences in Matrix Reasoning Are Linked to Functional Connectivity between Brain Regions Nominated by Parieto-Frontal Integration Theory.” Intelligence 87 (July): 101545. https://doi.org/10.1016/j.intell.2021.101545.
[28] Brochhagen T, Boleda G, Gualdoni E, Xu Y. (2023) From language development to language evolution: a unified view of human lexicon creativity.” Science. 381:431-436.

 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}