屏状核(claustrum)是成对位于大脑皮层第六层下方的薄片(图32.1)[1],接收来自皮层大部分区域的输入,并向皮层的大部分区域(图32.2)、包括内嗅皮层进行远程投射(图32.5D)[2,3],因此可能是大脑迭代的对抗生成网络(GAN)的关键环节之一(图32.2)。弗朗西斯·克里克博士在他人生的最后阶段对屏状核着迷,因为从与皮层区域的连接推测,它可能在整合意识感知方面发挥作用[4]。在整合多感官输入时,屏状核也发挥着作用[5]。我们在记忆的图15.3讲到,大脑皮层存储的信息更为丰富,覆盖各感官、运动和抽象概念(不认为只有前额叶负责认知,大部分事件都涉及多个区域),海马体可能已经主要是不断迭代的哈希索引(可以包含快速预览),而更小的屏状核更可能起到有判别的注意力的作用。这也不只是人工智能研究中常用的attention。
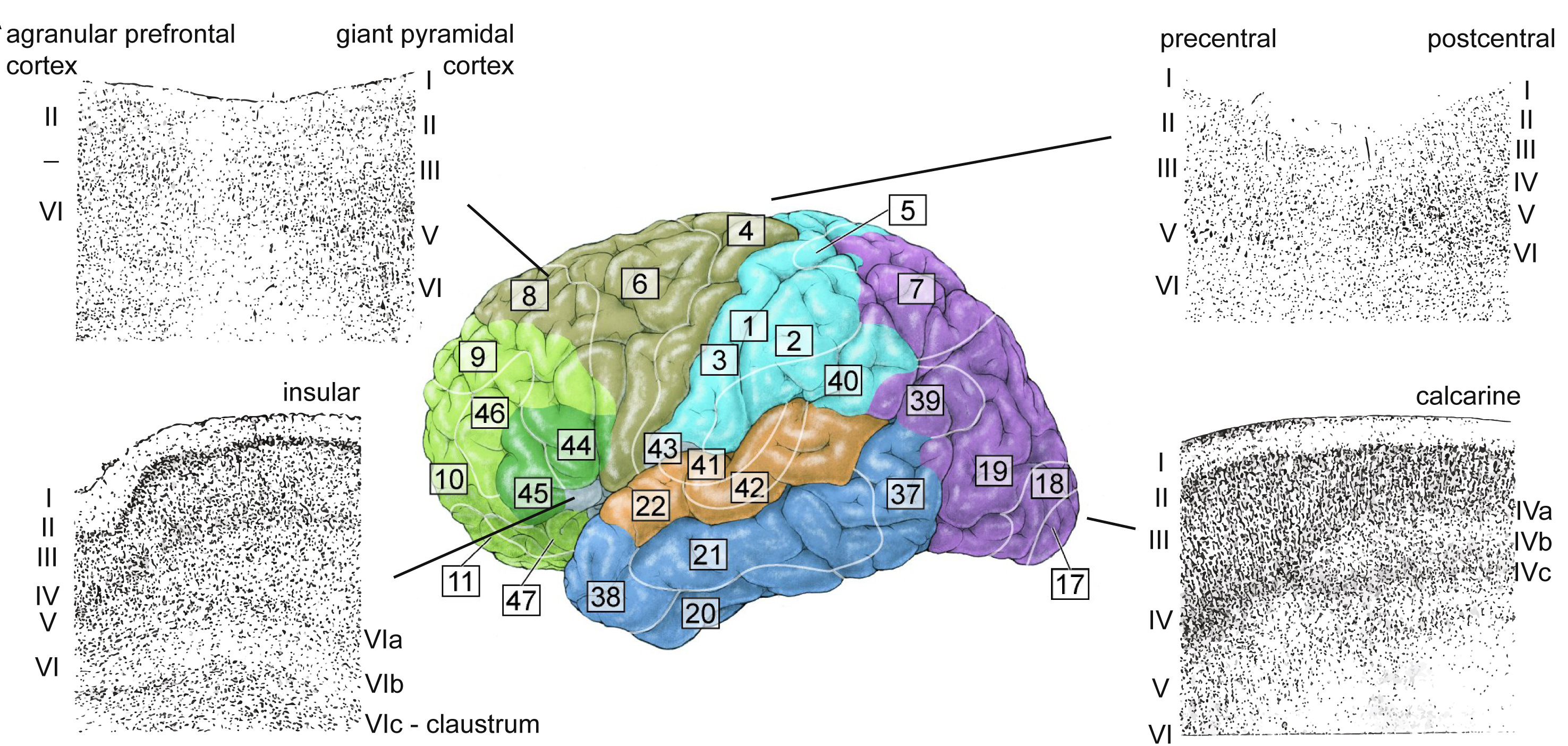
图32.1 图3.4 人脑的布罗德曼分区。神经元细胞核Nissl染色,可见I-VI层。屏状核位于岛叶皮层(Insular cortex)下后方白质内,与VIc亚层连续。内嗅皮层、海马等包在颞叶(Temporal lobe,深蓝色)里面,也看不到。紫色枕叶(Occipital lobe)在视觉皮层第IV层有分三亚层。来自[1],图3.4已有。
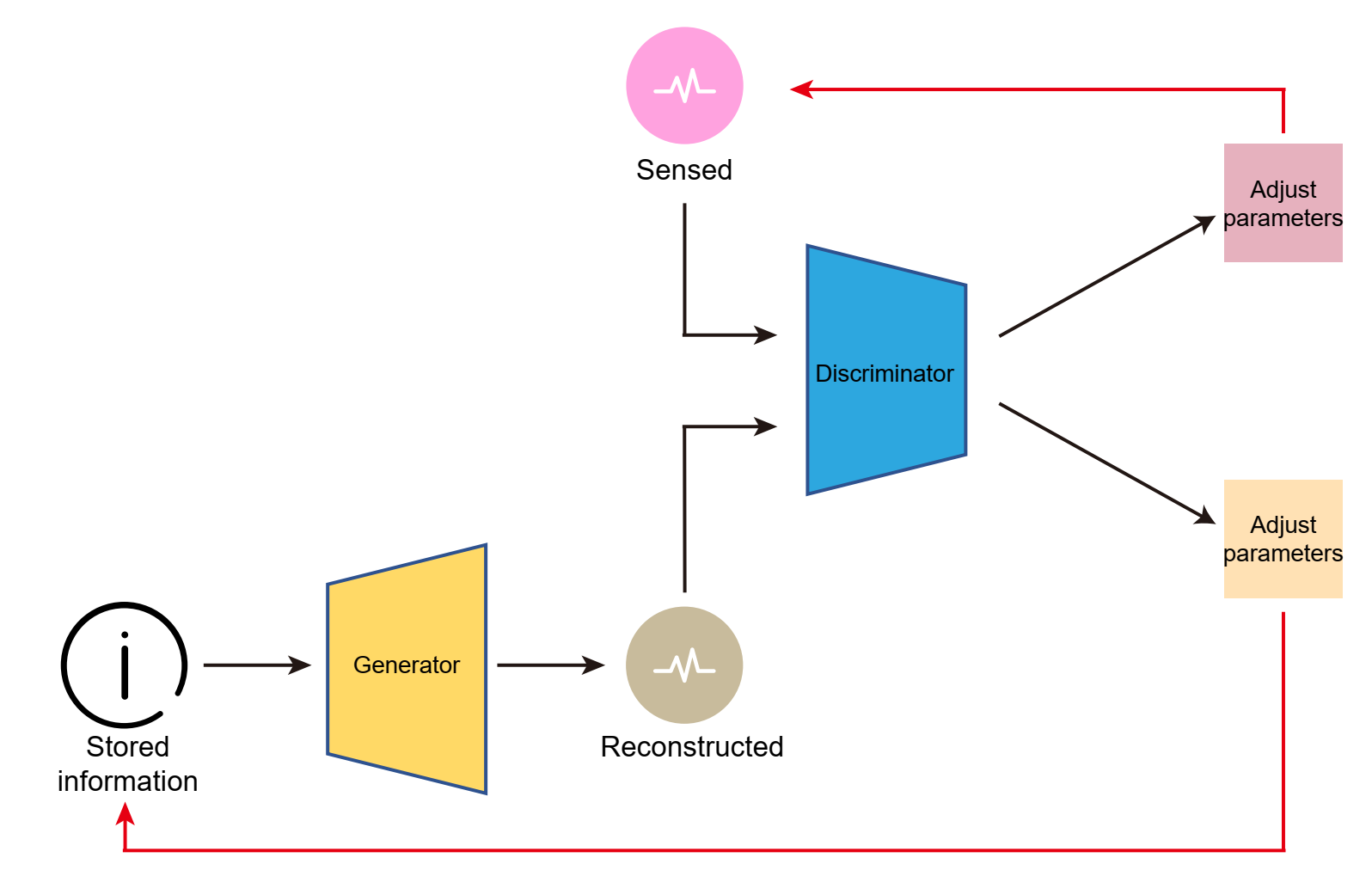
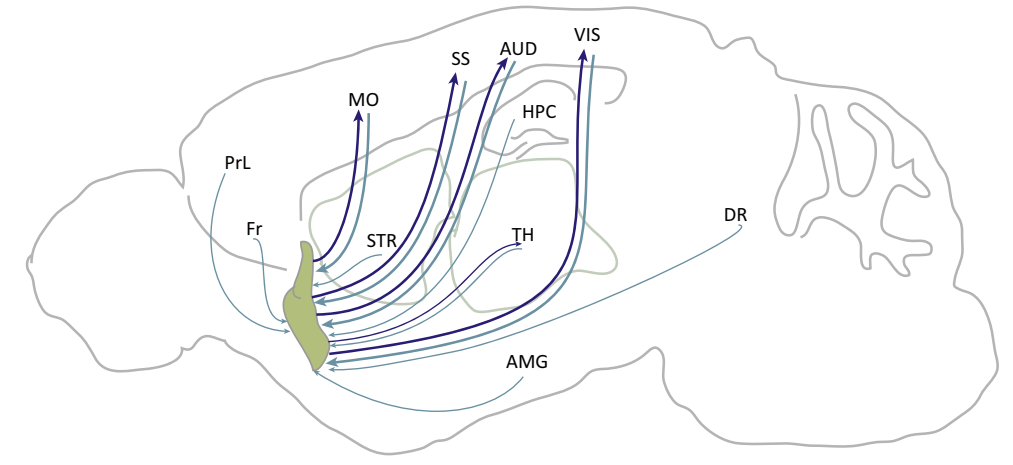
图32.2 (A) 在空间探索章节图27.4已提到的大脑的离线与在线学习兼备的对抗生成网络。(B)屏状核与大脑皮层各区域的互相投射。屏状核(绿色)的输入来自前额叶皮层以及来自纹状体、丘脑、杏仁核和背侧导管的亚皮质输入。几个非皮质(相对于皮质)区域从回旋带接收输入,包括梨状皮层、海马旁回和内嗅皮层,以及前额叶皮层的腹侧区域。回旋带主要从大脑皮层的第VI层接收输入,主要向第IV层锥体神经元或中间神经元和局部中间神经元发送输出,但也向第I/II层和第VI层发送输出。不同物种感觉系统的进化差异意味着同源区域并不总是存在,本图与大多数数据一致。PrL,前扣带皮层;ACA,前扣带区域;ILA,下扣带区域;MO,运动皮层;SS,体感皮层;ENT,内嗅皮层;人工智能,无颗粒岛叶区域;PIR,梨状区域;PrL,前扣带区域;Fr,前额叶皮层;AUD,听觉皮层;VIS,视觉皮层;STR,纹状体;TH,丘脑;AMG,杏仁核;HPC,海马;DR,背侧导管。在英文著作(https://www.jennystanford.com/9789814968782/neuroscience-for-artificial-intelligence/)已有,子图B来自[3]
在爬行动物胡须龙(Pogona vitticeps)中,也存在与屏状核类似的结构(虽然前文讲到,爬行类的大脑皮层可能对应尚未折叠到下方的海马皮层),在控制大脑状态方面发挥重要作用,并与慢波睡眠期间的尖波生成有关 [6],在另一项关于胡须龙的研究中,发现它两侧屏状核在慢波睡眠(NREM)期间各自产生尖波涟漪、并不协同;而在快速眼动睡眠REM中,两侧屏状核交替活动,一侧领先另一侧约 20 毫秒,在每段REM中仅切换顺序约一次(图32.2)[7]。两侧的屏状核并不直接竞争,而是通过中脑与后脑之间峡部的一个GABA核[7]。这种切换在哺乳动物大脑中是否也存在,有待研究。我们在REM的图22.1讲到了同侧海马皮层上背侧-腹侧theta波对应的从小到大位置场的切换(下一节讲更抽象的任务切换),如果能加上屏状核通过内嗅皮层推动的左右半球切换,将会是更完整的固有扫描机制。
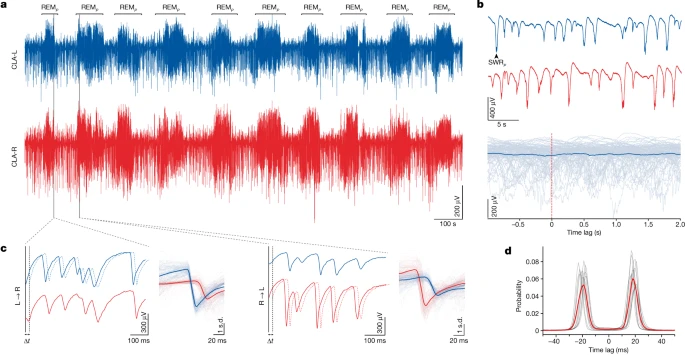
图32.3 胡须龙睡眠中屏状核的活动。(a) 慢波睡眠与REM交替中左侧(蓝)与右侧(红)屏状核的电活动。(b)上方,慢波睡眠时屏状核中尖波涟漪不同步;下方,假设一边受另一边激活的分析,蓝线为100次尖波涟漪平均。(c)子图a中两处REM的电活动时间轴放大,左图为左侧屏状核领先于右侧屏状核,右图已切换为右侧屏状核领先为左侧屏状核。(d)REM中左右关联的平均峰(红线),在正、负约20ms各有一峰。共12晚,7只动物,9小时睡眠,每只记录22-26万次去极化(single sharp negative extracellular potential, SN)。来自[7]。
从大脑皮层到屏状核的双向输入-输出连接可能是前文提到的GAN中的鉴别器部分(图32.2),左右两侧的连接还有差异(图32.5)[3,8,9,10,17]。屏状核-MEC的回放/预放(规划)可能更有助于矢量导航或计划,而皮层-海马位置细胞的回放可能更有助于记忆和准确定位。虽然屏状核在大脑皮层灰质中只占约0.25%,但在大鼠中屏状核对运动皮层、感官皮层的投射都已经至少有数十万个神经元[11],与皮层各区域都有足够的交互。
虽然没有监测屏状核,大鼠的迷宫实验清晰展示了MEC-海马的左右切换(图32.4)[12],在前进中“左顾右盼”,在头脑中作有规律的探索,很好地覆盖了行进路线周围的区域。这种切换在快速眼动睡眠时仍然存在[12]。虽然大鼠实验传统上为了严格设计,都只是简单的迷宫。不难想象人类的运动,比如跳水、自由式小轮车等开始实施前也需要预放一步步动作。现场环境的干扰相当于占用了内存(下一节讨论的工作记忆),影响了预放的整体协同。
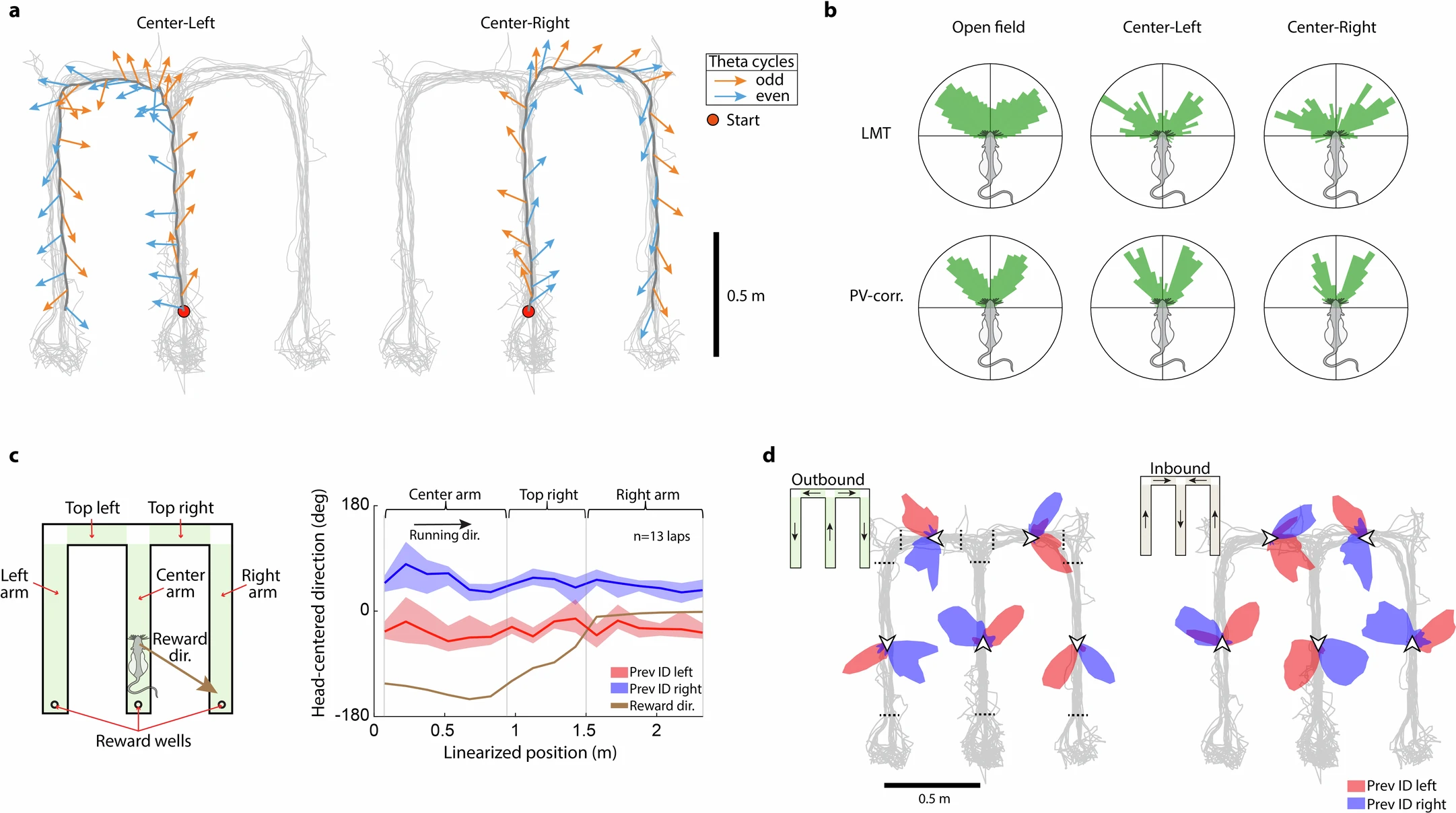
图32.4 大鼠在迷宫中有目标地探索时头脑内部方向的左右切换。(a)从红点出发,在迷宫中向左或向右探索目标,循环的theta波的左右(奇偶)箭头显示为不同颜色。(b)极坐标直方图展示两种不同方法(上方,Latent manifold tuning,LMT,下方,头部朝向调制曲线)得到的在旷场、迷宫向左或向右时的内部方向分布。(c)内部方向与当前奖励的位置无关。LMT得出的内部方向红或蓝表示。(d)极坐标直方图在迷宫具体位置的展示,红蓝与子图c一致。来自[12]
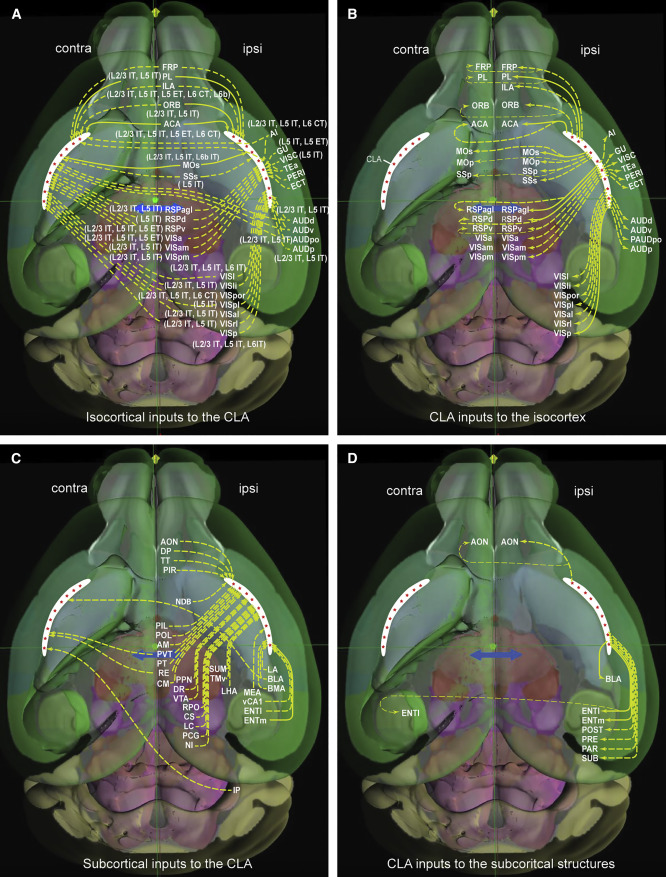
图32.5 小鼠左右屏状核与大脑皮层和皮层下区域的连接。白色表示屏状核,红点代表锥体神经元(principal neurons),文中分为9组。(以右侧为准,)contra(lateral)即对侧投射,ipsi(lateral)即同侧投射。黄色箭头实现代表强投射,虚线代表弱投射。(A)等皮层至屏状核。主要有intra-telencephalic (IT) neurons in layer 2 (L2) to L6, extra-telencephalic (ET) neurons in L5, and corticothalamic (CT) neurons in L6。(B)屏状核至等皮层。与图32.2类似。(C)皮层下区域至屏状核。EPd, basolateral amygdala nucleus (BLA), paraventricular nucleus (PVT), central medial nucleus (CM), dorsal raphe nucleus (DR) 。(D)屏状核至皮层下区域。ENTl,lateral entorhinal area; ENTm, medial entorhinal area。除了强投射至内嗅皮层,还弱投射至海马周边,postsubiculum (POST), presubiculum (PRE), parasubiculum (PAR)。BLA,基底神经节。来自[17]
Dehaene等人在1998年提出了大脑中的全局工作空间理论[13](图32.6)。在原始版本中,神经元以自上而下的方式进行调控与组织,而笔者倾向于认为它们更接近于自下而上的方式。例如,前文提到的嗅觉,记忆涟漪和预演;但来自ACC-MEC的注意力可以作为自上而下的信号。现在,随着分布式计算与联邦学习的相关知识普及,可能更容易理解这种分布式全局工作空间。每个模块显示其当前状态,这些状态可能由重大事件协调和更新。神经元间的低频放电被用来维持工作记忆[14]。
全局工作空间可以说是一种并行计算与串行计算相结合的工作模式。无论涉及何种感官,全局协调可能都利用了对应的高级脑区域(例如,视觉区域用于做梦;梨状皮层用于嗅觉空间表示[15])。人们可以推测,用于全局协调的神经元的工作空间的大小和主导模块可能因个体和生理状态而异。工作记忆,包括大脑状态间的有序转换,受到前额叶皮层多巴胺的调节,并且在精神分裂症患者中出现工作记忆受损[16]。我们在下一章会进一步从图论角度推测海马体的分区数目。
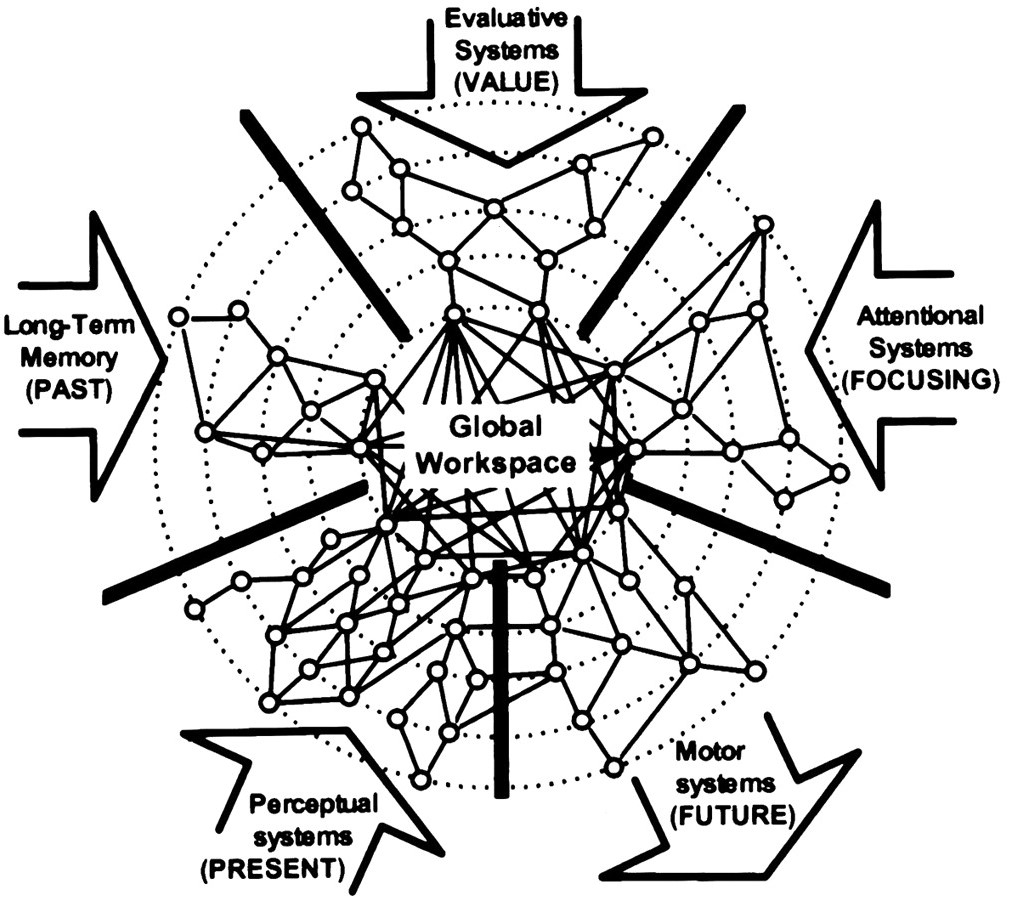
图32.6 全局工作空间中五种主要处理模块的示意图。来自[13]
参考文献:
[1] Cadwell, C. R., Bhaduri, A., Mostajo-Radji, M. A., Keefe, M. G. & Nowakowski, T. J. Development and arealization of the cerebral cortex. Neuron 103, 980–1004 (2019).
[2] Kitanishi, Takuma, and Naoki Matsuo. 2017. “Organization of the Claustrum-to-Entorhinal Cortical Connection in Mice.” The Journal of Neuroscience : The Official Journal of the Society for Neuroscience 37 (2): 269–80. https://doi.org/10.1523/JNEUROSCI.1360-16.2016.
[3] Goll, Yael, Gal Atlan, and Ami Citri. 2015. “Attention: The Claustrum.” Trends in Neurosciences 38 (8): 486–95. https://doi.org/10.1016/j.tins.2015.05.006.
[4] Crick, F. C. & Koch, C. What is the function of the claustrum? Philos. Trans. R. Soc. Lond. B. Biol. Sci. 360, 1271–9 (2005).
[5] Remedios, R., Logothetis, N. K., & Kayser, C. (2010). Unimodal responses prevail within the multisensory claustrum. The Journal of Neuroscience : The Official Journal of the Society for Neuroscience, 30(39), 12902–12907. https://doi.org/10.1523/JNEUROSCI.2937-10.2010
[6] Norimoto, H., Fenk, L. A., Li, H.-H., Tosches, M. A., Gallego-Flores, T., Hain, D., Reiter, S., Kobayashi, R., Macias, A., Arends, A., Klinkmann, M., & Laurent, G. (2020). A claustrum in reptiles and its role in slow-wave sleep. Nature, 578(7795), 413–418. https://doi.org/10.1038/s41586-020-1993-6
[7] Fenk, L. A., Riquelme, J. L., & Laurent, G. (2023). Interhemispheric competition during sleep. Nature, 616(7956), 312–318. https://doi.org/10.1038/s41586-023-05827-w
[8] Kitanishi, Takuma, and Naoki Matsuo. 2017. “Organization of the Claustrum-to-Entorhinal Cortical Connection in Mice.” The Journal of Neuroscience : The Official Journal of the Society for Neuroscience 37 (2): 269–80. https://doi.org/10.1523/JNEUROSCI.1360-16.2016.
[9] Kitanishi, Takuma, and Naoki Matsuo. 2017. “Organization of the Claustrum-to-Entorhinal Cortical Connection in Mice.” The Journal of Neuroscience : The Official Journal of the Society for Neuroscience 37 (2): 269–80. https://doi.org/10.1523/JNEUROSCI.1360-16.2016.
[10] Peng, Hanchuan, Peng Xie, Lijuan Liu, Xiuli Kuang, Yimin Wang, Lei Qu, Hui Gong, et al. 2021. “Morphological Diversity of Single Neurons in Molecularly Defined Cell Types.” Nature 598 (7879): 174–81. https://doi.org/10.1038/s41586-021-03941-1.
[11] Kowiański P, Moryś J, Sadowski M, Dziewiatkowski J. (2000). Qualitative and quantitative differences in the motor and somatosensory cortical projections of the rat claustrum--combined retrograde transport and stereological studies. Folia Morphol (Warsz). 59(2):111-119.
[12] Vollan, A.Z., Gardner, R.J., Moser, MB. et al. Left–right-alternating theta sweeps in entorhinal–hippocampal maps of space. Nature 639, 995–1005 (2025). https://doi.org/10.1038/s41586-024-08527-1
[13] Dehaene, S., M. Kerszberg, and J.-P. Changeux. 1998. “A Neuronal Model of a Global Workspace in Effortful Cognitive Tasks.” Proceedings of the National Academy of Sciences 95 (24): 14529–34. https://doi.org/10.1073/pnas.95.24.14529.
[14] S, Kornblith, Quian Quiroga R, Koch C, Fried I, and Mormann F. 2017. “Persistent Single-Neuron Activity during Working Memory in the Human Medial Temporal Lobe.” Current Biology : CB 27 (7): 1026–32. https://doi.org/10.1016/J.CUB.2017.02.013.
[15] Poo, Cindy, Gautam Agarwal, Niccolò Bonacchi, and Zachary F. Mainen. 2022. “Spatial Maps in Piriform Cortex during Olfactory Navigation.” Nature 601 (7894): 595–99. https://doi.org/10.1038/s41586-021-04242-3.
[16] Braun, Urs, Anais Harneit, Giulio Pergola, Tommaso Menara, Axel Schäfer, Richard F Betzel, Zhenxiang Zang, et al. 2021. “Brain Network Dynamics during Working Memory Are Modulated by Dopamine and Diminished in Schizophrenia.” Nature Communications 12 (1): 3478. https://doi.org/10.1038/s41467-021-23694-9.
[17] Wang Q, Wang Y, Kuo HC, Xie P, Kuang X, Hirokawa KE, Naeemi M, Yao S, Mallory M, Ouellette B, Lesnar P, Li Y, Ye M, Chen C, Xiong W, Ahmadinia L, El-Hifnawi L, Cetin A, Sorensen SA, Harris JA, Zeng H, Koch C. Regional and cell-type-specific afferent and efferent projections of the mouse claustrum. Cell Rep. 2023 Feb 28;42(2):112118. doi: 10.1016/j.celrep.2023.112118.

0
推荐
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}