阅读:0
听报道
我们在前面章节已经提到,在大鼠、小鼠中研究最多的空间探索,还可以被用于社交探索等其他功能,语言等功能可能也涉及屏状核-内嗅皮层-海马体与大脑皮层的交互。进一步推演,逻辑推理等抽象活动可能也是在一个个小平面上展开的。很少有动物既有足够的脑细胞,又不必把主要精力用于获得温饱和传宗接代。
路径图(path diagram)(例如图37.1)作为因果推断和反事实的工具[1,2],其命名恰当地反映了其与空间导航的相似性。现代科研论文结果的起承转合,也是一个一维导航、排除其他可能性的过程(图37.2)。每个事件都是一个节点,就像每个事件都可通过大脑皮层与海马体的相互作用被存储与提取复用一样。有些节点比其他节点更容易被激活(包括幻想某人叫自己的名字[3],幻听)。更短、更通用的路径将更高效、更具泛化能力,节省细胞(例如网格细胞相对于位置细胞,图27.2;右海马“位置细胞”用于在故事中的确定事件间的边界[4]),并在每次成功运行后积累更高的权重。这类似强化学习中的内在奖励。
合适的归因可以是一个逐步完善、细化的过程。早年欧洲的船长和水手们只知道柠檬可以预防坏血病,那就可能提供了维生素C含量低的品种,甚至提供了煮过的酸饮料。当还是出现了大批坏血病时,就有不相信这一招的了(神经元释放乙酰胆碱来调整参数,强化学习的第31节提到[5],REM睡眠图)。
人类独特的地方在于,从口口相传到使用文字记载,知识会系统地从我们素未谋面的人那里传递下来,这样我们就可以或由点到线,或自上而下地积累有用的模式(例如,几百年前某人的生活),超越单个动物所能经历的时空尺度。然而人类能够进行因果推理,这并非是因为形而上的原因,或是由于某种灵丹妙药。正如Judea Pearl所倡导的那样,大脑有一种内生的倾向展开因果推断[1,2]。
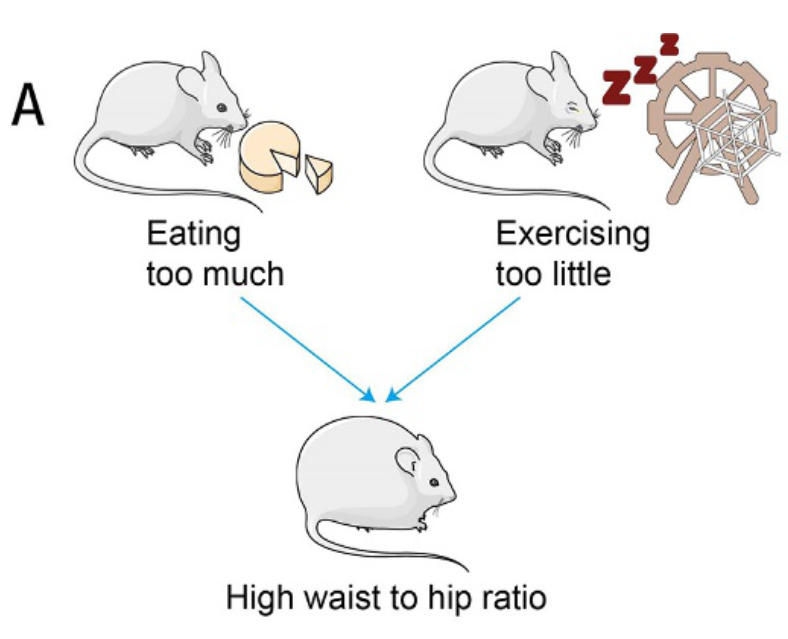
图37.1 因果路径图示例。这是一个对撞(collider)的例子,即两条路径指向同一个结果。此时不能统计处理使结果恒定,否则会使两个原因呈现负相关的假象。来自著作www.sciencedirect.com/book/monograph/9780323913690/investigating-human-diseases-with-the-microbiome第6章
本节从空间探索的角度探讨人和动物可以,但人工智能还做不到的第三层因果证据--反事实,即如果没有A,是否还会到B。第一层因果证据即关联,第二层称为干预,例如随机临床试验。
因果关系不可逆是狭义相对论的基本假设之一。量子纠缠作为微观粒子的特性,也没有泛化到宏观事件。人脑在学习这些论述时,既涉及新细胞对新概念的表征,也涉及已有存储的更新和拓展。前文还讲过,新生突触倾向于赫布学习(关联),成熟突触倾向于反赫布学习(区分)。
在现有可能路径的框架中(图37.1,图37.2,图37.3),提出一个反事实问题,相当于询问当在心智地图上禁用一条路径时,是否仍能到达目标位置。该问题的答案,在确认其它可能路径存在后,还与替代路径(如贝叶斯概率分配[1,2])和禁用路径在地图上当前的权重(指导选择模式和各路径现有概率的先验信息)有关(图37.1,图37.3)。
20世纪下半叶以来盛行的分子生物学研究,也未尝不是一种从一个蛋白质到另一个蛋白质的有向图的构建活动。已报道的通路可以迁移过来作为补全,而新增的关键路径需要更严谨地测试(图37.2)。例如用不同方法证明同一段结论,已排除特定方法可能造成的假象。
当AI可以从多篇文献中整理出逻辑框架时,科学杂志的编辑工作就可以完全自动化。已知和未知形成一个巨大的网格,大部分人只需要结论及其可信度。每个结论(节点)对应了一系列证据,对猜想的建立(创造性连接)或对结论证实的贡献也将是科研评价体系摆脱杂志名称和杂志论文平均引用数(所谓影响因子)的重要契机。
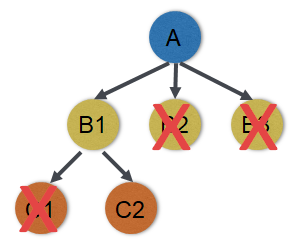
图37.2 现代科研论文通常每篇确立一个结论,论述逻辑大致如图所示,需要排除其他可能性。A通过B导致C也是科研经费申请书常用的套路。经常被提及的奥卡姆剃刀原则,在著作www.sciencedirect.com/book/monograph/9780323913690/investigating-human-diseases-with-the-microbiome第6章也有转述E.T.Jaynes教授从贝叶斯的概率分布角度的解释[2]
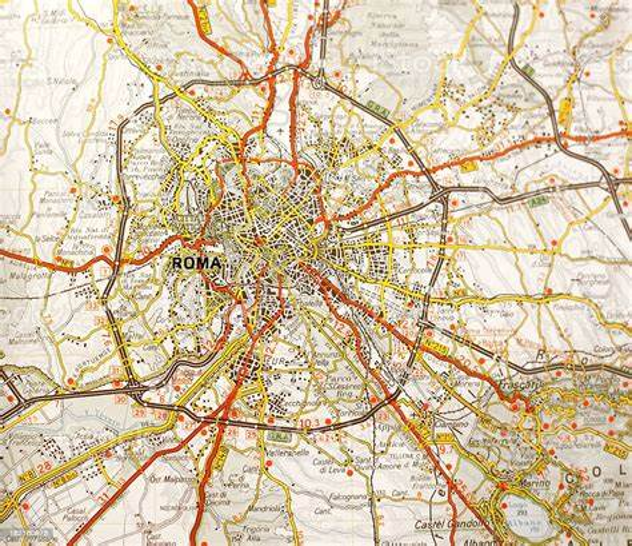
图37.3 罗马地图。路有粗细不同。目标导航可能开始也只需要大致正确,离得近了再细化具体要到哪个地点。反事实推断意味着假设移除一条路径,并估计我们是否仍然能够到达目标地点。根据我们先前的知识,如果只有一条主要路径,那么关于因果关系,反事实问题就不存在了。每个人都有他或她自己的心理地图,但常识知识在大多数人被存储在短路径中,具有较大的权重。来源:https://www.istockphoto.com/photo/all-roads-lead-to-rome-the-map-gm182160878-19950566.
基于空间探索的因果路径图还可以解释阴性结果为什么不如阳性结果那么容易被大家接受与传播。一个信息量较大的实验可以导致概率的重新分配。而阳性结果建立的路径上,概率分布得以集中。阴性结果没有提出新路径,而是试图使已有的路径变细,释放出来的概率没有落实分配。从神经元树突棘的角度,一旦变粗就很难再变细(第17节),可能需要小胶质细胞等直接吞噬。
为大脑投入能量是一项昂贵的投资。没有多少物种能够奢侈地去思考那些与生存无关的事情[6],晚一些成熟,也可能对大脑有利[7]。
小鼠大脑皮层中兴奋性锥体神经元的树突棘阵列在树突轴上间距约1 μm,每个突触宽约0.3 μm[8];海马体中锥体神经元的突触更大[9];大篮状细胞等抑制性神经元突触作用于细胞体时的突触也较大(图17.5)。
Braitenberg 和Schüz通过对小鼠大脑电子显微镜图像的长期研究估计,大脑皮层中的任意两个锥体神经元都可以由两个或三个突触连接起来的[8],这是大脑皮层兼顾稀疏性与穷举可能性的基础设置之一。大脑皮层的锥体神经元可能存储具体感官、动作(脊髓、小脑等配合),人、事、物与概念,由此相连组成的所有可能关联,会是一个天文数字,也是人脑进行因果关系推断的生物学基础。这样的小世界网络,意味着任意两个概念之间,要将其连起来,所需的中间步骤也就2到3个(屏状核-MEC网格-海马可以整体调用既成范式)。当我们说出自己的推理时,拆分后是不是也就是两到三步了?
新皮质不同层,连同丘脑和屏状核的神经元,构成了许多可以调整的连接[10,11]。这些连接组合,构成了用以发现因果关系的素材,在确定为因果关系之前,相关性可先在大脑中以中间神经元调制的弱连接存储。创造性的首次连接也可以非常有价值。
中间神经元到锥体神经元的轴突输出可能经常是多对多的(37.4)[12,13]。一个有趣的可能性是,两个概念之间的弱联系可能通过放松从一个中间神经元到多个锥体神经元的平行抑制(锥体神经元也可能有多个轴突输出),即同时激活来实现。如果从VIP中间神经元抑制多个其他中间神经元开始,这组关联的锥体神经元的数量和距离都可以相当可观。这也是赫布学习可能的细胞基础之一。如果经常关联出现的概念距离太远,可能还是在更合适的地方另存一份较好(睡眠整理),MEC-海马对该范式的调用也可以更高效。
而因果关系更可能是确定的从一个锥体神经元连接到下一个锥体神经元的通路,可视为在两个概念间建立了有向的路径。这种调用的效率应该是比关联关系更高的。大脑神经活动对刺激物的反应,从剧烈的、孩子气的大范围振荡,逐渐精进为反应更迅速的、更模块化反复调用的模式。但因果连接相比相关连接,更加难以被重写,过多的因果连接对于网络的可塑性可能是有代价的。更多信息事物的刺激可能有助于大脑维持一个没有过度拟合、过度细分的神经元网络。而目前人工智能算法甚至经常没有抑制性中间神经元。
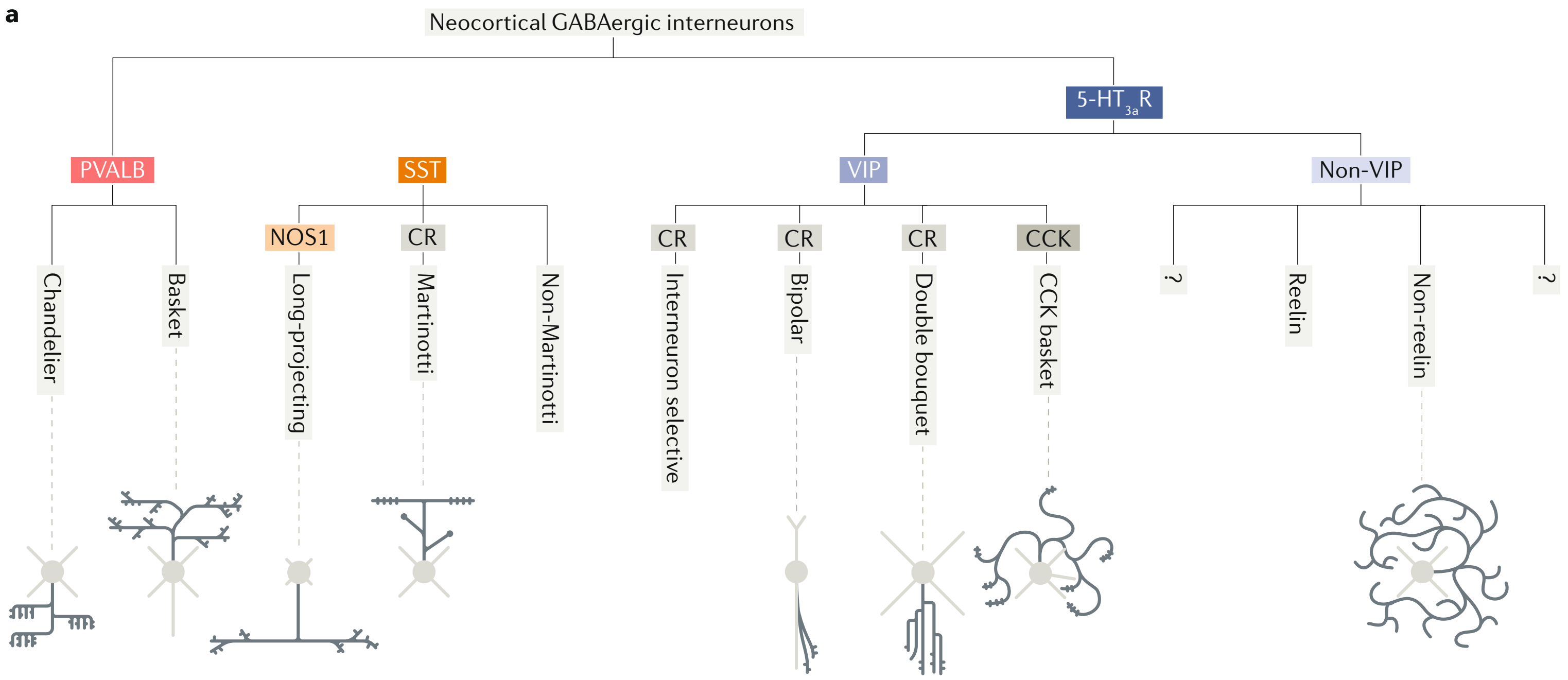
图37.4 几种主要的抑制性中间神经元在轴突存在多输出。大类与图5.2、图7.3、图21.2一致。人脑可能有更多缺乏研究的细分类群。浅灰色为树突,深灰色为轴突。来自[12]
作为拥挤大脑中的初始设置,抑制性中间神经元、星形胶质细胞(astrocytes,第14节,第19节)等可能在锥体神经元连接到其它锥体神经元之前,占据其突触表面,阻止不必要连接的诞生。这些抑制性神经元在合适的信息刺激后才释放被保护的锥体神经元,让它们开始建立点到点的路径。从这个角度考虑,一旦建立起了单向连接的因果关系,再要修改它们也是有困难的,尤其是连接的概率过大,在突触间表现得过于粗壮时。这是网络时代经常讨论的对谣言的坚信,以及“杠精”的神经机制。
我们在上几节讲语言时就提到,海马-MEC网格可能被投射到适当的新皮层,上述投射搜索现有的感官和概念信息(例如嗅觉探索[14]),并在心智模拟中形成规划或框架(图15.3,图32.2)。有了一个或多个锚点后,将可构成一个具有更大灵活性的迁移学习,根据搜索过程中出现的存储模式对现有表征进行补全,并将其组合成一个新的序列。
通过对已存储的网格的迁移,神经系统在下一次使用这些神经元时,对可能的连接和事件(点和线)存在先验预期,有助于预测后续步骤。各类规则以向量形式(可能反映了theta波等脑电波的走向,第26节)存储,还伴随着规则周边的信息。经历每个新事件,大脑都会与现有的模式集进行哈希索引。如果事件与大脑表征相一致,那就是令人满意的,并且更有可能在未来被再次使用(更大的树突棘不容易变得更大)。
由于网格式存储模式,导致新事件与已有存储的差异处于30°时,网格细胞的激活程度低,在这个角度上外部和现有模式的对齐可能不会导致神经元的调整(图37.5)。而且网格中每个节点的强弱也有差异,从戒备较低的节点逐步切入,然后再反转,可能有更强的效果。这使得记忆更新存在一个阈值,只有在阈值之上才可以参与更新,包括海马CA3-CA1的概率更新、变动更大的从海马齿状回加入新神经元细节等(第16节)。除了神经元生长时间不足,这可能是某些人的认知模型尽管经过了进一步的训练,与现实的拟合缺乏改进的一个潜在解释,尤其是在社交等抽象领域。
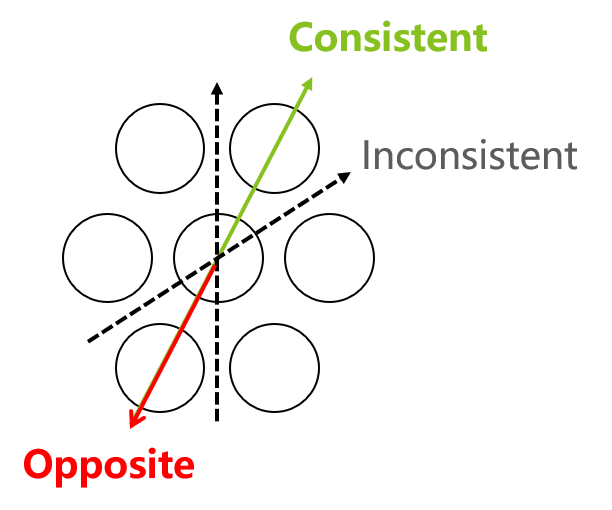
图37.5 一个用于判断新事件相对于大脑中现有向量的一致性的六边形网格。这些圆圈可以代表皮层中的一个或多个神经元细胞。与空间导航期间的网格细胞活动类似,实箭头表示完美对齐,而虚箭头表示30°角度时的最低活动。与现有向量相反的事件应引起神经元的强烈活动。英文著作https://www.jennystanford.com/9789814968782/neuroscience-for-artificial-intelligence/已有
对于意外的结果,更大规模的神经活动会被触发,以招募能够编码新颖性的树突棘、树突和神经元。在教学和演讲中,引用熟悉的例子再展示意外情况是常用的技巧。设问以后留出合适的时间,让听众的大脑运行一下,但又得及时接着讲。熟悉性可能提供了用于对比和信息存储的参考点,而意外性则产生了强烈的信号(图37.5),增加了招募新突触、新分支和新细胞的可能性。事件由此根据意外程度被给出优先级,以便于在无需大量的训练数据时,一个完整的画面得以迅速出现(尽管存在不确定性)。类似的方式,可用以启发少尝试学习(few shot learning)的算法架构设计。
在前文中,我们讨论了事件之间的“距离”(例如,在新地点使用海马体中现有的连接模式)和回放(强化和探索各种可能性)/预放(计划)。基于这些经验,在遇到新事件事可能可以预留出一个合适尺度的区域,以便后续填充细节。从大网格开始,使用一个或多个小网格对现有模式进行细化。海马体的索引词和来自多个类别的感官输入也可以帮助定义位置。如果没有网格框架,从已知点出发,推演的步骤越多,误差就会越大。
在来自其他地方[15,16]的输入作用下,单个神经元中的巧合检测可能是在因果图中加入依赖多事件的、更复杂的条件概率。同一树突分支上的相邻多个突触(可能来自具有延迟的并行电路)甚至单个树突棘可能都参与上述条件概率的更新(图37.6)。存储在大脑皮层不同区域的范式也可能被调用到同一个神经元中来进行新的计算。
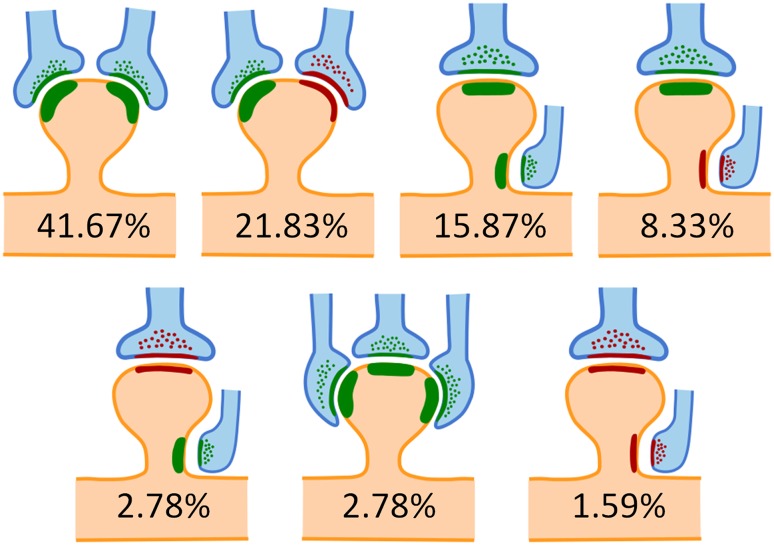
图37.6 未成年大鼠感觉皮层各层统计归纳的树突棘接受多个输入的情形。观察的4558个树突棘中5.57% (254个) 有2-3个突触,各自占比如图所示,红色为兴奋性突触、绿色为抑制性突触。还有约5%的更稀有的组合没有画出。来自[17],图17.4复用
大脑会从常识中习得有用的规则,而不是被填鸭式的投喂。大脑在填补认知缺失(探索未知),并为未来积累有用的模式时,会产生内生奖励,而这可能是好奇心的本质所在。在此过程中,反馈哈希模型被用于搜索和记忆信息,每晚经由睡眠对新收集的信息进行整理,这导致比现有算法更加可迭代和自适应的GAN。在快速搜索后,之前学到的稀疏匹配模式可能通过扣带回-内嗅皮层-海马波投射到大脑皮层,以指导新场景下的行为。因此,对于成年人来说,永远不会面对完全陌生的新情况,但与经历过的模式都松散匹配,及其新组合可能会鼓励更多的即兴创作。
当一个先前成功的模式被用于当前任务,大脑的GAN在调用生成(图32.2)时,应当以什么样的采样频率检测大脑中现有的预测与外部事实的差异?当同时存在第二个或第三个模式在排队时,资源分配如何随时间变化?大脑的左右屏状核,海马前后推进、对应大小网格的theta波等在这些方面的参数是否最优,是有趣的研究方向。
每个大脑中的心智地图都不可避免地有所不同,但仍然可以在不同大脑间达成共识。心智地图中被忽略的路径可能会成为通往重要事物的门户,有时这被称为灵感。
试探性关联,其结果可能会缺乏详细路径(机制),但大脑进行的贝叶斯运算会迅速在树突棘中积累每个已生效规则的权重,从而构建更精细的地图。当干预措施(如随机对照试验)试图建立路径时,反事实推断在心理上阻止对应路径的产生,并询问我们是否仍可以到达目的地。心智地图存储在每个人的大脑中,因此如果我们无法到达那个地方,我们就知道。这可能是反事实对于大脑来说很容易的神经基础。但请不要拒绝新的关联。
参考文献:
[1] Pearl, Judea, and Dana Mackenzie. 2018. The Book of Why.
[2] Jaynes, E. T. 2003. Probability Theory: The Logic of Science.
[3] Horga, Guillermo, and Anissa Abi-Dargham. 2019. “An Integrative Framework for Perceptual Disturbances in Psychosis.” Nature Reviews Neuroscience 20 (12): 763–78. https://doi.org/10.1038/s41583-019-0234-1.
[4] Cohn-Sheehy, Brendan I., Angelique I. Delarazan, Zachariah M. Reagh, Jordan E. Crivelli-Decker, Kamin Kim, Alexander J. Barnett, Jeffrey M. Zacks, and Charan Ranganath. 2021. “The Hippocampus Constructs Narrative Memories across Distant Events.” Current Biology 31 (22): 4935-4945.e7. https://doi.org/10.1016/j.cub.2021.09.013.
[5] Chantranupong, L., Beron, C.C., Zimmer, J.A. et al. Dopamine and glutamate regulate striatal acetylcholine in decision-making. Nature 621, 577–585 (2023). https://doi.org/10.1038/s41586-023-06492-9
[6] Kraft, Thomas S., Vivek V. Venkataraman, Ian J. Wallace, Alyssa N. Crittenden, Nicholas B. Holowka, Jonathan Stieglitz, Jacob Harris, et al. 2021. “The Energetics of Uniquely Human Subsistence Strategies.” Science 374 (6575). https://doi.org/10.1126/science.abf0130.
[7] Tooley, Ursula A., Danielle S. Bassett, and Allyson P. Mackey. 2021. “Environmental Influences on the Pace of Brain Development.” Nature Reviews Neuroscience 22 (6): 372–384. https://doi.org/10.1038/s41583-021-00457-5.
[8] Braitenberg, V. & Schüz, A. Cortex: statistics and geometry of neuronal connectivity. Cortex Stat. Geom. Neuronal Connect. (1998) doi:10.1007/978-3-662-03733-1.
[9] Padmanabhan, Pranesh, Andrew Kneynsberg, and Jürgen Götz. 2021. “Super-Resolution Microscopy: A Closer Look at Synaptic Dysfunction in Alzheimer Disease.” Nature Reviews Neuroscience 22 (12): 723–40. https://doi.org/10.1038/s41583-021-00531-y.
[10] Shepherd, Gordon M.G., and Naoki Yamawaki. 2021. “Untangling the Cortico-Thalamo-Cortical Loop: Cellular Pieces of a Knotty Circuit Puzzle.” Nature Reviews Neuroscience 22 (7): 389–406. https://doi.org/10.1038/s41583-021-00459-3.
[11] Larkum, Matthew. 2013. “A Cellular Mechanism for Cortical Associations: An Organizing Principle for the Cerebral Cortex.” Trends in Neurosciences 36 (3): 141–51. https://doi.org/10.1016/j.tins.2012.11.006.
[12] Huang, Z. J. & Paul, A. The diversity of GABAergic neurons and neural communication elements. Nat. Rev. Neurosci. 20, 563–572 (2019).
[13] Hage, Travis A, Alice Bosma-Moody, Christopher A Baker, Megan B Kratz, Luke Campagnola, Tim Jarsky, Hongkui Zeng, and Gabe J Murphy. (2022). “Synaptic Connectivity to L2/3 of Primary Visual Cortex Measured by Two-Photon Optogenetic Stimulation.” eLife 11. https://doi.org/10.7554/eLife.71103.
[14] Bao, Xiaojun, Eva Gjorgieva, Laura K Shanahan, James D Howard, Thorsten Kahnt, and Jay A Gottfried. 2019. “Grid-like Neural Representations Support Olfactory Navigation of a Two-Dimensional Odor Space.” Neuron 102 (5): 1066-1075.e5. https://doi.org/10.1016/j.neuron.2019.03.034.
[15] Larkum, Matthew. 2013. “A Cellular Mechanism for Cortical Associations: An Organizing Principle for the Cerebral Cortex.” Trends in Neurosciences 36 (3): 141–51. https://doi.org/10.1016/j.tins.2012.11.006.
[16] Egger, Robert, Rajeevan T. Narayanan, Jason M. Guest, Arco Bast, Daniel Udvary, Luis F. Messore, Suman Das, Christiaan P.J. de Kock, and Marcel Oberlaender. 2020. “Cortical Output Is Gated by Horizontally Projecting Neurons in the Deep Layers.” Neuron 105 (1): 122-137.e8. https://doi.org/10.1016/J.NEURON.2019.10.011.
[17] Santuy, A., Rodriguez, J. R., DeFelipe, J. & Merchan-Perez, A. Volume electron microscopy of the distribution of synapses in the neuropil of the juvenile rat somatosensory cortex. Brain Struct. Funct. 223, 77–90 (2018).
话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}