阅读:0
听报道
非侵入式脑电(EEG)检测到从亚赫兹到上百赫兹的脑电波(睡眠章节详述),最快的这一两百赫兹的一类就被称为尖波涟漪(Sharp-wave ripples,图18.1)[1]。可能因为背景更干净,也可能因为泡在脑脊液中确实信号更强、更同步,尖波涟漪在慢波睡眠(非快速眼动睡眠NREM的较深阶段)中相比清醒时更明显。对记忆回放(replay)和尖波涟漪(sharp wave ripples),在目前的大量文献中也缺乏落实到神经元物理基础的理解。因为小鼠研究通常涉及让小鼠在线性轨道上跑步,关于记忆回放的理解,还经常会说回放得比实际跑的时候快了。笔者认为关键问题是记忆回放过程涉及了多少突触。数十到数百毫秒的自发回放[2],在脑电波表现为尖波涟漪[3,4]。
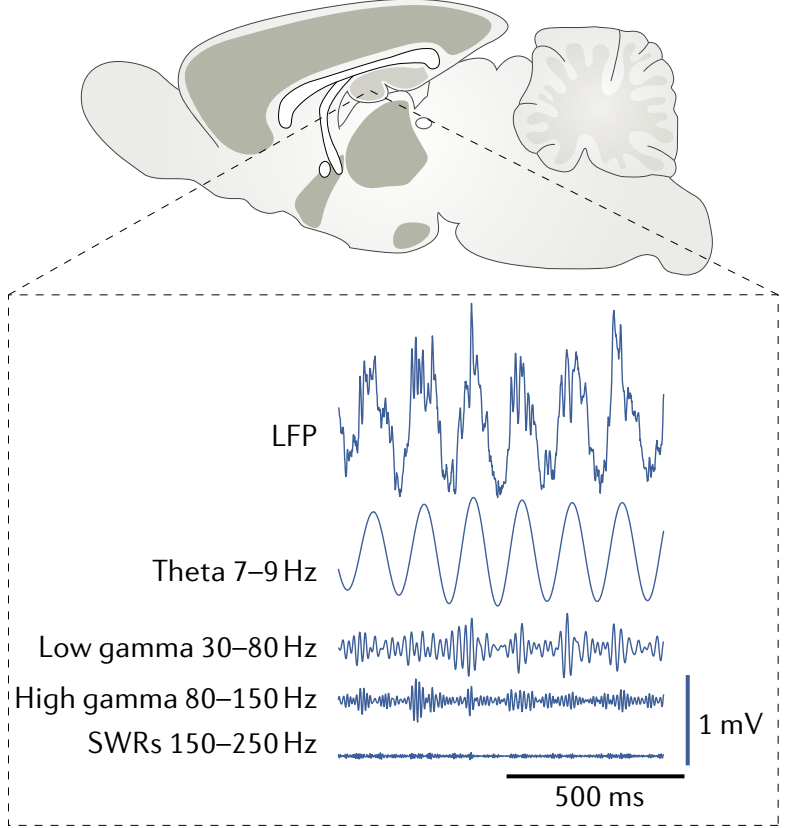
图18.1 小鼠快速眼动睡眠(REM)时海马体按频率划分的几种脑电波示意图。除了LFP(local field potentials)为原始记录外,拆解出的几种波都经过了过滤。来自[1]
相比更虚无缥缈的理解,尖波涟漪的约200Hz的频率实际上与单个突触的电活动所需时间相一致。虽然不同类型的神经元略有差异,一个突触放电通常历时5-10毫秒,皮层锥体神经元的兴奋性突触约5毫秒 [5,6,7] ;钙离子从一个树突棘到树突轴的扩散可以持续15 毫秒[5] ,这段时间可能激活与同一树突轴相连的其它细胞的突触,以及神经回路内其他细胞,然后这些细胞产生各自的电流和钙离子释放。这样整个过程可以持续上百毫秒,就与成簇的尖波涟漪一致了,是一个具备物质基础的可能的解释。上述的突触间电信号传递与树突棘到树突轴的钙离子传递也可能包括同时并行发生的、就如同增加了绕路(经过其它神经元)但前往同一终点神经元的神经回路。
电刺激小鼠海马CA1区域的神经元,可以在平均23毫秒后激活前扣带皮层(Anterior cingulate cortex,ACC)的神经元(5-60ms 范围)[8],包括了两组神经元各自需要的时间。海马CA1尖波涟漪活动峰值与前扣带皮层 I型神经元放电峰值之间的平均延迟期为12ms (6ー32ms)[8]。与之相一致,从前扣带皮层到海马的神经连接通常是单一突触的[9]。该实验刺激下,大约四分之一的 CA1尖波涟漪事件导致了前扣带皮i 型神经元的激活,两者的电活动并不完全耦合。这是神经元间信号传递方面的过滤与稀疏化(睡眠的正则化作用后面章节再讲)。
由于神经元的树突棘阵列中包含了所有必要的信息,记忆的回放并不需要激活每一个突触,相反,每一簇树突棘中最先去极化(可以导致动作电位)的突触可能具有足够的代表性,也就是我们大脑中一闪而过、但是足以知道是什么事的记忆回放(图18.2、图18.3)。这是既稀疏又省电的模式,不是非要整个包含细节地重复一遍。
这与当前类脑计算和认知神经学中的回放机制不同。当前大多数尝试只是通过回放来减少灾难性遗忘。亚细胞层面的回放将不仅用于巩固记忆,而是可以通过激活部分突触进行组合计算,产生新的,可能在进化上更有利的表征[10],可能称为“复盘”更合适。我们在介绍空间探索、强化学习的时候再细讲往前放(计划)与往后放(回放,或者说复盘)。
基于回放的表征学习,类似于以自监督的形式从文本学到词向量,但生物大脑并不是以更契合上下文为目标优化的。每个新增的记忆都会经历稀疏化的过程,只要后续提取记忆时这些特征够用,也就不需要推倒重来,可能略有调整即可以更优化。
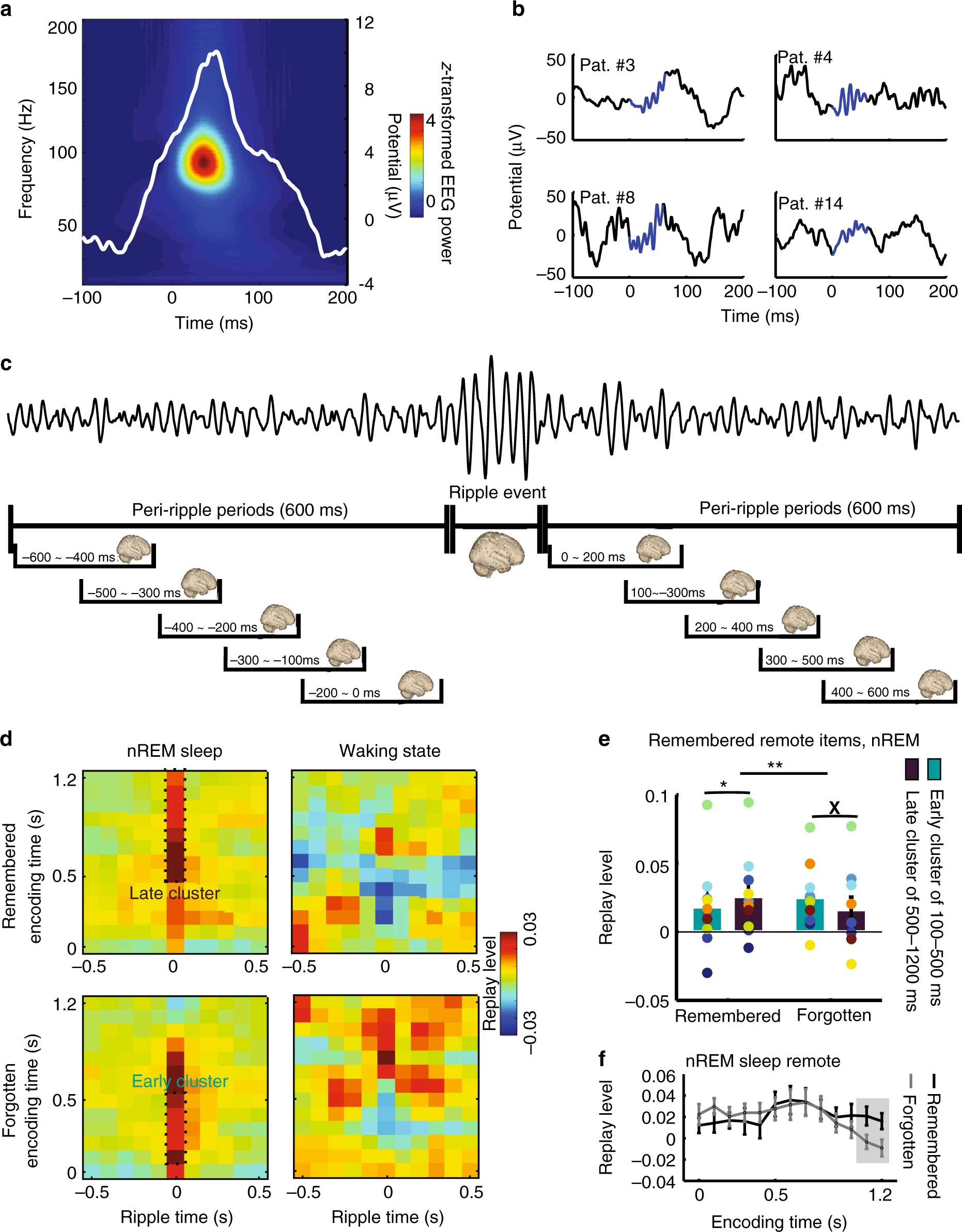
图18.2 尖波涟漪代表的记忆回放。对12例癫痫患者分别植入脑电图(EEG)电极进行诊断。在午睡1小时前学习80张风景或建筑物的照片,在午睡后又学习80张照片,后一组没有经过睡眠期间的回放。然后这些患者志愿者需要判断测试中展示的照片(已学习的80+80张再混入新的80张)是否看过。(A) 颅内EEG检测到的海马体电活动,展示尖波涟漪前后的数百毫秒时间段。数据经40Hz以上过滤。(B) 几位患者的尖波涟漪事件(类似图18.1,有其他波叠加)。(C)尖波涟漪前后的电活动展示,尖波事件后可以分为早发簇(100-500 ms)和晚发簇(500-1200 ms)。(D)NREM睡眠时的晚发簇与早发簇。(E,F)NREM时晚发簇与成功的图片记忆有关,早发簇与没记住有关。来自[11]
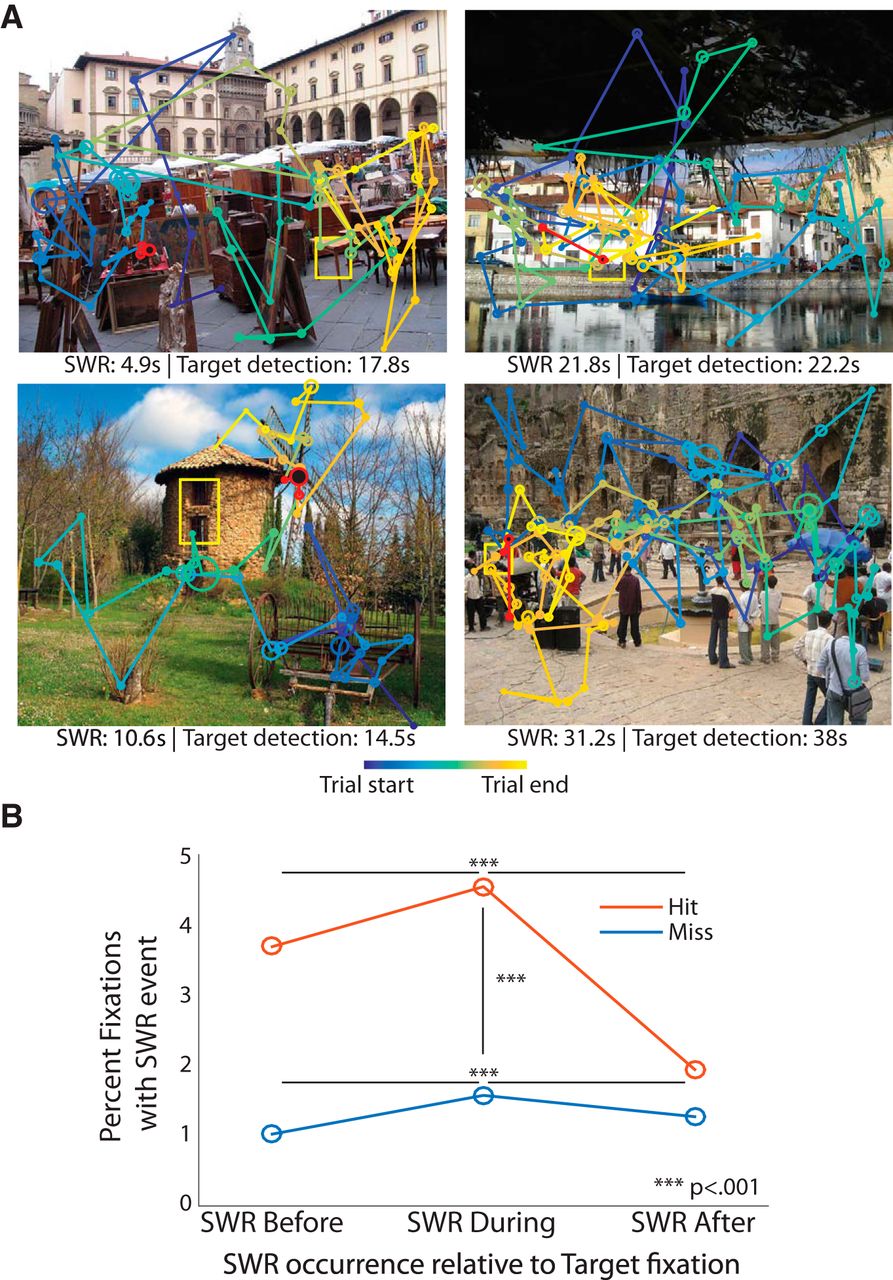
图18.3 受试猴在图片中找红色光点对应的尖波涟漪。共4只猴在海马CA3/DG 区域被植入了电极。(A)图片上展示了4次找红点的眼动轨迹。海马体尖波涟漪略早于或与找到同时。(B)统计了尖波涟漪与找到红点的关系。来自[12]
由于无法对健康人直接进行细胞层面的检测(只有少量因医疗原因植入电极的癫痫患者可以),故当前研究只能对健康人通过非侵入式的EEG检测脑区的电波,对脑波仅按照频率作了分类命名,而缺乏更物理层面的描述。在一项癫痫患者图像记忆的研究中,午睡非快速眼动睡眠(NREM)期间的尖波涟漪回放与成功的图片辨认有关(图18.2)[11]。相对较晚的回放可能对应了新特征的学习,相比更早、更熟练的回放,辨认效果更好(图18.2)。而一项恒河猴在图片中找红色光点的研究更直观展示了尖波涟漪对应的搜索成功(图18.3)[12]。
小鼠在一个首次接触的场地上跑了几次后,一系列位置记忆可能就需要新引入一些树突棘、树突轴和神经元(空间探索的章节经常会看到这些所谓位置细胞,place cells)。我们在图16.1已经讲到,海马CA1的神经元主要由内嗅皮层和海马CA3输入,激活产生新的表征。在记忆的表征引入了新的神经元(不一定是新生的,可以只是调用进入小网络)后,尖波涟漪的放电随之被优化(图18.4)[13]。这可能是对新引入神经元的必要性的一个评估,后续还可以进一步优化神经元之间的分配。
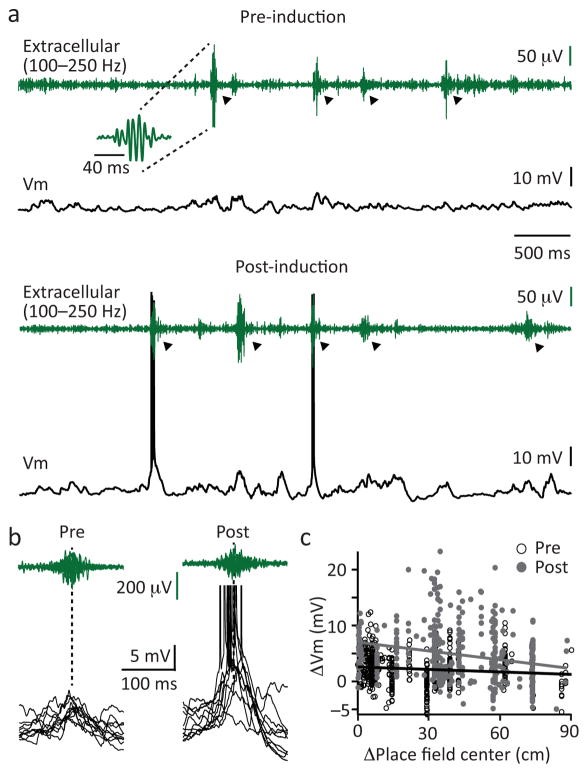
图18.4 小鼠海马CA1新增位置表征后,尖波涟漪伴随的去极化(突触前神经元的Vm升高,离子通道更可能打开)和动作电位。(a)新增位置细胞前后的尖波涟漪和去极化。(b)新增位置细胞前后的连续动作电位。(c)新增位置细胞表征后,尖波涟漪伴随的未触发动作电位的去极化 (∆Vm) 幅度在新增表征的中心更高(灰点拟合的灰线);而新增前的表征更平缓(空心点拟合的黑线)。来自[13]
除了经典的自细胞体输出的轴突,近期研究还发现小鼠海马CA1相当多的锥体神经元含有从细胞体周边树突发端的轴突,使CA1锥体神经元参与同层网络化(手拉手的感觉)的尖波涟漪活动[14]。海马CA1、CA3的初始设置都包含涉及较多神经元的自发性尖波涟漪振荡,而被分配给明确的线索后,海马体锥体神经元就不再参与这些自发的尖波涟漪[15]。同一项研究对CA3的模拟(8000个兴奋性的锥体神经元、150个抑制性的中间神经元)还观察到了回放,涉及特定位置的有序激活[15]。
综上所述,在本书第15节介绍的海马体-大脑皮层的哈希索引反馈迭代的记忆模型中,尖波涟漪代表的突触放电体现了精确到神经元及其树突棘的海马体-大脑皮层信息提取。
在睡眠章节会进一步解释的一些较慢的波,如 delta 波(由中间神经元导致的同步) 、theta 波,则可能对应了更大范围的搜索过程,在这些更大尺度下,尖波涟漪代表了搜索的结果。而上一节讲到,树突棘因使用而健壮,因此尖波涟漪是突触们竞争性激活的体现,反映了神经元网络特性相对于初始均衡状态的偏离。然后我们就该讲睡眠了。
参考文献:
[1] Adamantidis, A. R., Gutierrez Herrera, C. & Gent, T. C. Oscillating circuitries in the sleeping brain. Nat. Rev. Neurosci. 20, 746–762 (2019).
[2] Ólafsdóttir, H. Freyja, Daniel Bush, and Caswell Barry. 2018. “The Role of Hippocampal Replay in Memory and Planning.” Current Biology : CB 28 (1): R37–50. https://doi.org/10.1016/j.cub.2017.10.073.
[3] Krause, Emma L., and Jan Drugowitsch. 2022. “A Large Majority of Awake Hippocampal Sharp-Wave Ripples Feature Spatial Trajectories with Momentum.” Neuron 110 (4): 722-733.e8. https://doi.org/10.1016/j.neuron.2021.11.014.
[4] Valero, M., Zutshi, I., Yoon, E. & Buzsáki, G. Probing subthreshold dynamics of hippocampal neurons by pulsed optogenetics. Science (80-.). 375, 570–574 (2022).
[5] Noguchi, Jun, Masanori Matsuzaki, Graham C.R. Ellis-Davies, and Haruo Kasai. 2005. “Spine-Neck Geometry Determines NMDA Receptor-Dependent Ca2+ Signaling in Dendrites.” Neuron 46 (4): 609–22. https://doi.org/10.1016/J.NEURON.2005.03.015.
[6] Braitenberg, Valentino, and Almut Schüz. 1998. Cortex: Statistics and Geometry of Neuronal Connectivity. Cortex: Statistics and Geometry of Neuronal Connectivity. Berlin, Heidelberg: Springer Berlin Heidelberg. https://doi.org/10.1007/978-3-662-03733-1.
[7] Tripathy, Shreejoy J., Shawn D. Burton, Matthew Geramita, Richard C. Gerkin, and Nathaniel N. Urban. 2015. “Brain-Wide Analysis of Electrophysiological Diversity Yields Novel Categorization of Mammalian Neuron Types.” Journal of Neurophysiology 113 (10): 3474–89. https://doi.org/10.1152/JN.00237.2015.
[8] Wang, Dong V, and Satoshi Ikemoto. 2016. “Coordinated Interaction between Hippocampal Sharp-Wave Ripples and Anterior Cingulate Unit Activity.” The Journal of Neuroscience : The Official Journal of the Society for Neuroscience 36 (41): 10663–72. https://doi.org/10.1523/JNEUROSCI.1042-16.2016.
[9] Rajasethupathy, Priyamvada, Sethuraman Sankaran, James H Marshel, Christina K Kim, Emily Ferenczi, Soo Yeun Lee, Andre Berndt, et al. 2015. “Projections from Neocortex Mediate Top-down Control of Memory Retrieval.” Nature 526 (7575): 653–59. https://doi.org/10.1038/nature15389.
[10] Kurth-Nelson, Z., Behrens, T., Wayne, G., Miller, K., Luettgau, L., Dolan, R., Liu, Y., & Schwartenbeck, P. (2023). Replay and compositional computation. Neuron, 111(4), 454–469. https://doi.org/10.1016/j.neuron.2022.12.028
[11] Zhang, Hui, Juergen Fell, and Nikolai Axmacher. 2018. “Electrophysiological Mechanisms of Human Memory Consolidation.” Nature Communications 9 (1): 4103. https://doi.org/10.1038/s41467-018-06553-y.
[12] Leonard, T. K. et al. Sharp wave ripples during visual exploration in the primate hippocampus. J. Neurosci. 35, 14771–14782 (2015).
[13] Bittner, K. C. et al. Conjunctive input processing drives feature selectivity in hippocampal CA1 neurons. Nat. Neurosci. 18, 1133–1142 (2015).
[14] Hodapp, A., Kaiser, M. E. & Thome, C., et al. Dendritic axon origin enables information gating by perisomatic inhibition in pyramidal neurons. Science 377 (6613), 1448–1452 (2022).
[14] Hodapp, A., Kaiser, M. E. & Thome, C., et al. Dendritic axon origin enables information gating by perisomatic inhibition in pyramidal neurons. Science 377 (6613), 1448–1452 (2022).
[15] Terada, S. et al. Adaptive stimulus selection for consolidation in the hippocampus. Nature 601, 240–244 (2022).
话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}