如果不能准确地估计方向、距离和位置,对动物来说意味着丧失关键机会,例如食物、潜在配偶。在前文简要提到过的海马体位置细胞(place cells,图25.1A)的基础上,本节介绍位于内侧内嗅皮层(MEC,人后内侧内嗅皮层,pmEC, posteromedial entorhinal cortex)的网格细胞(grid cells,图25.1B)。与上一节所谓time cells类似,这些细胞的命名也是根据大鼠或小鼠在场地上活动时神经元的激活情况来定义的。这种脱离细胞基础的定义使得名称泛滥,也使得大量不呈现典型六边形网格的神经元不被叫作网格细胞、甚至另有名称(比如band cells,下一节图)。超过10% 的MEC神经元完整显示出六边形网格状放电模式[1,2],被称为网格细胞。
即使在一个开放的空间,网格细胞活跃的点阵也是相对于感知的场地边界倾斜的(图25.1B),这样每个位置相比不倾斜状况下更加独特(对于圆形边界,这是或许是个问题,但也可以偏离中心放置),经过几个细胞的组合可能就可以对场地内任意一个点唯一确定坐标。
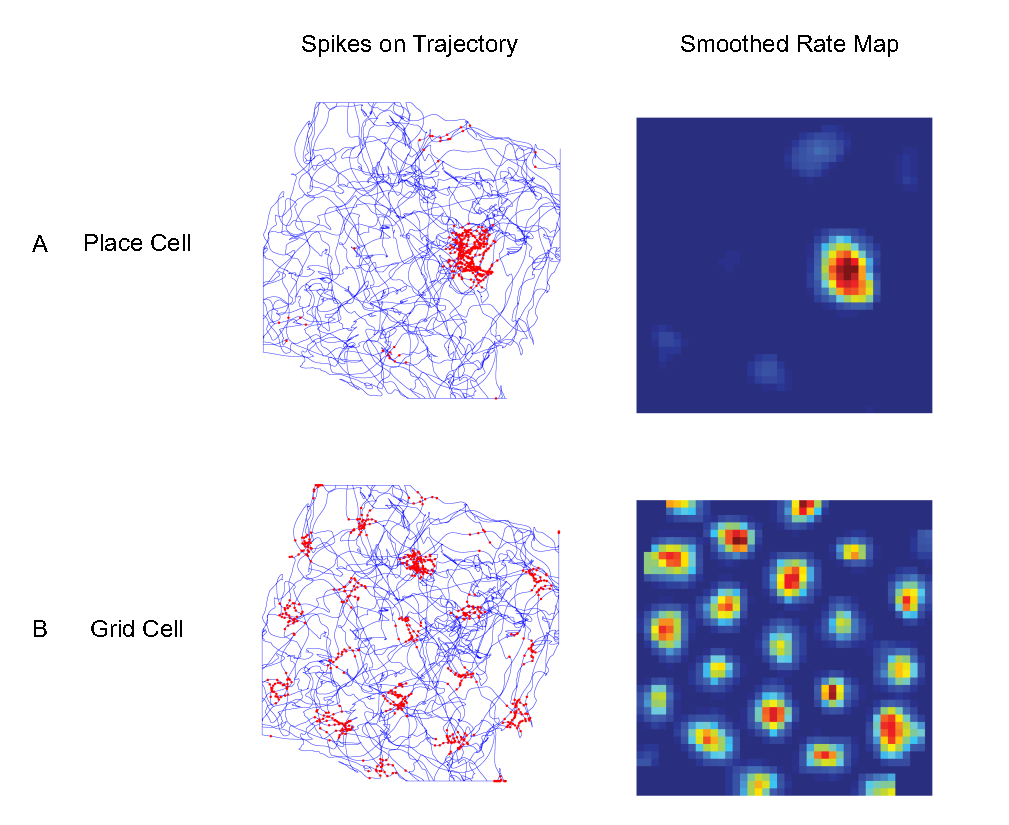
图25.1 (A)海马体单个位置细胞在旷场实验时的放电场。(B)内侧内嗅皮层单个网格细胞(我们下一节会看到这涉及很多细胞)的放电场。第一列显示了插有电极的动物在试验过程中对应轨迹(蓝线)上的细胞放电位置(红点)。第二列显示了平滑处理后细胞的放电率图,从蓝色(最小值)到红色(最大值)。虽然大多数位置细胞的放电位置只有一个(较大,且有梯度),但网格细胞的放电场形成一个周期性的三角形矩阵,覆盖动物可达的整个环境。
与位置野(place field,即位置细胞在场地中放电的区域)从小鼠或大鼠海马背侧到腹侧逐渐增大的现象类似(在人类中为从后侧到前侧,这也是θ波相位传播的方向,图22.1),研究发现网格细胞的网格尺寸也沿背侧到腹侧轴逐渐增大,并且每个网格偏好的方向有所不同[3,4,5]。
放电点阵更密的网格细胞,其数量相对更多;而在腹侧端,对应最大尺度的网格细胞似乎与对应较小尺度的细胞共存(图25.2A,B)[6]。我们在后一节推测它们的神经元基础。
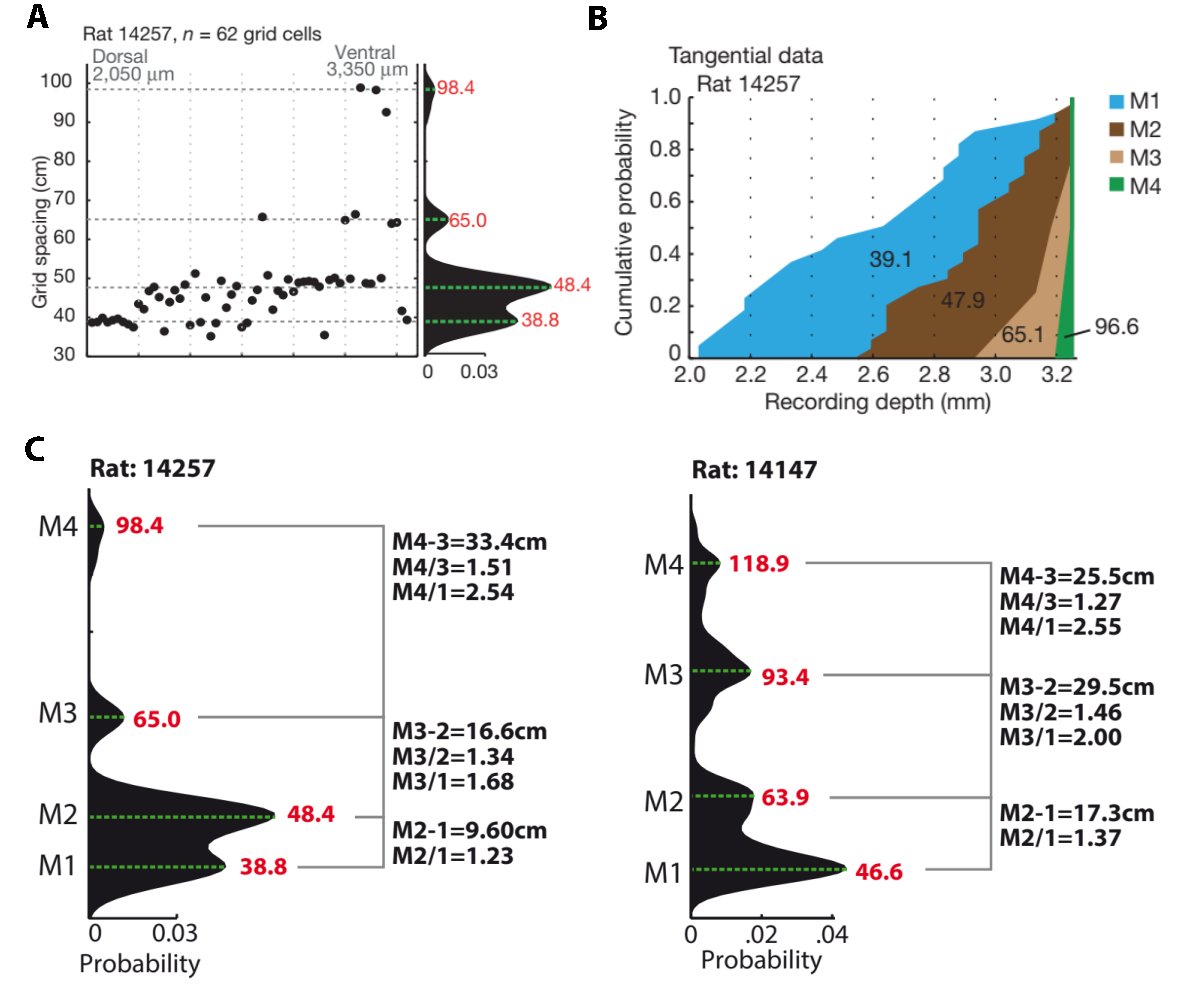
图25.2 网格模块被组织成重叠的带状结构。(A)单个大鼠在背腹轴上的不同网格细胞放电格子大小。点表示单个细胞。细胞按顺序排列,以避免在相似深度处的细胞重叠。右侧,分布的核平滑密度(KSD)估计,得出各模块的格子大小(红字,以厘米为单位)。(B)作为背腹位置函数的网格模块累积分布。x轴为插入电极深度。各模块内的平均网格间距,与A接近。(C)左、右两只大鼠的四个网格模块的间距和比例关系。左边与A、B是同一只大鼠。来自[6]
目前研究显示大鼠的网格细胞可以被聚类成约五个模块[6, 7]。有趣的是,其中最小的网格似乎相当于大鼠体长的两倍(图25.2A,C),而最大的网格可能还受限于实验场地。仅图25.2C的这两只大鼠已经体现出网格细胞大小的差异,扯得远些这甚至可以表现为“性格”。
与理论计算一致,相邻网格细胞对应区域的大小之比不超过1.5,可以嵌套确定坐标(图25.3)[8,9]。设想一下,在一个陌生的街区,知道要去的方向,即较大的矢量,而具体路径则需要较小的网格来适应一些七拐八拐(后面还会讲到网格细胞与位置细胞的配合,进一步提高定位的准确性)。而当网格细胞簇之间的间隔超过一个或两个模块时,两簇网格细胞对应的尺度比率(相除后)会更大(图25.2C),这可能导致在定义位置时出现灾难性错误,在缺少中间尺度的网格细胞作指导的时候产生随机性[8,9]。这在新环境中探索时可能很有用。
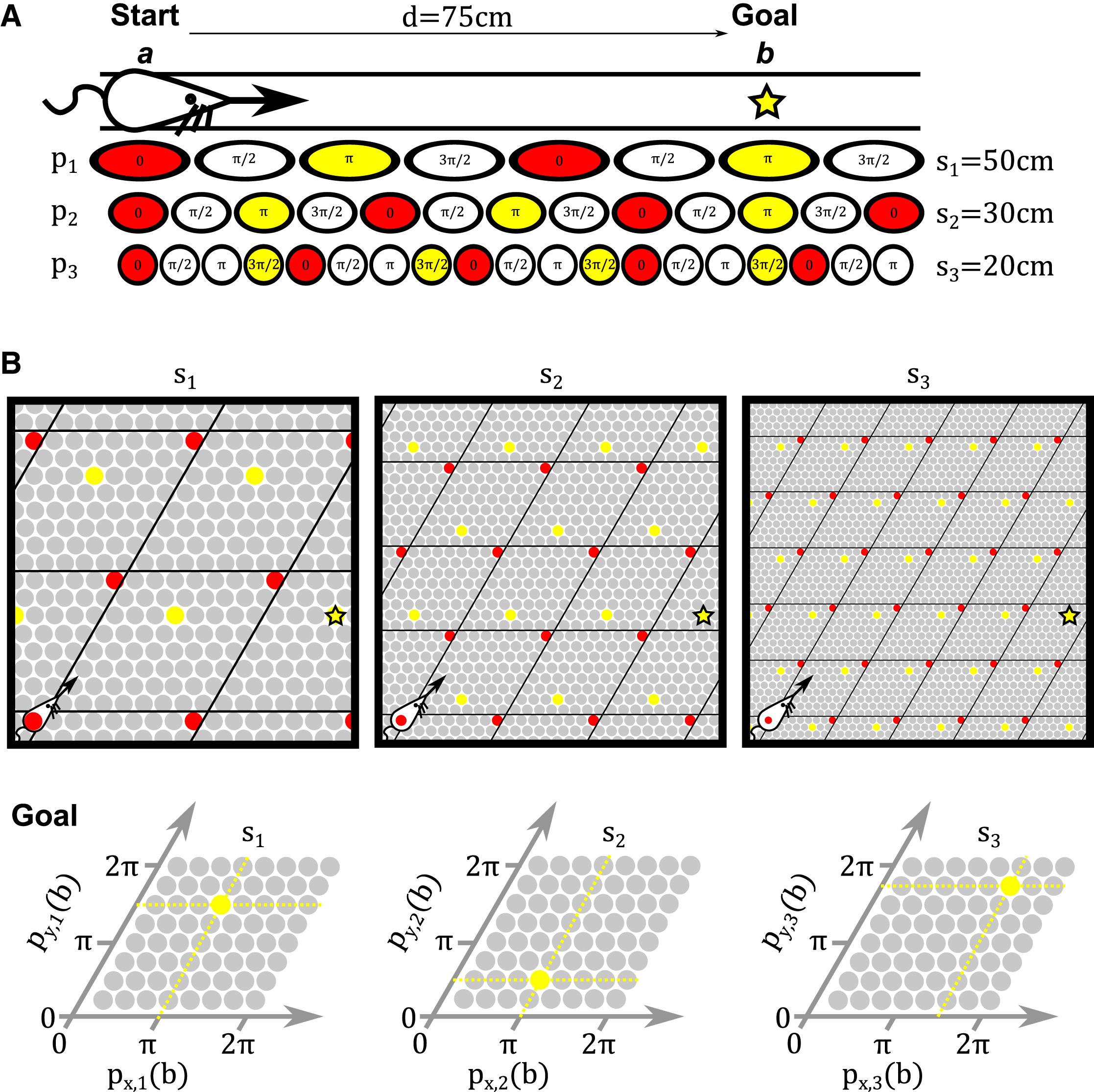
图25.3 向量空间中导航问题可以通过不同大小的网格来解决。(A) 在一维空间中,我们可以将每个网格细胞模块可视化为一个环,支持一个以相位π为中心的一组细胞活动。起始位置a(红色)和目标位置b(黄色)之间的位移d可以使用这些位置的网格细胞表示中计算出来。(B) 在二维空间中,我们可以将每个网格细胞模块可视化为一个扭曲的环面(twisted torus),支持一个以相位π→=(px,i,py,i)为中心的单个活动峰,沿着网格图案的单位“瓦片”的主轴(即单位向量x→和y→)。起始位置和目标位置之间的距离和方向可以通过这些位置的网格细胞表示来定义。来自[9]
动物的网格大小分布受限于发育过程中的梯度(下一节),工程上更自由的网格组合是不是可以实现什么新功能呢。
参考文献:
[1] Rueckemann, Jon W, Marielena Sosa, Lisa M Giocomo, and Elizabeth A Buffalo. 2021. “The Grid Code for Ordered Experience.” Nature Reviews. Neuroscience 22 (10): 637–49. https://doi.org/10.1038/s41583-021-00499-9.
[2] Hardcastle, Kiah, Niru Maheswaranathan, Surya Ganguli, and Lisa M Giocomo. 2017. “A Multiplexed, Heterogeneous, and Adaptive Code for Navigation in Medial Entorhinal Cortex.” Neuron 94 (2): 375-387.e7.
[3] Hafting, Torkel, Marianne Fyhn, Tora Bonnevie, May-Britt Moser, and Edvard I Moser. 2008. “Hippocampus-Independent Phase Precession in Entorhinal Grid Cells.” Nature 453 (7199): 1248–52. https://doi.org/10.1038/nature06957.
[4] Guanella, Alexis, Daniel Kiper, and Paul Verschure. 2007. “A Model of Grid Cells Based on a Twisted Torus Topology.” International Journal of Neural Systems 17 (4): 231–40. https://doi.org/10.1142/S0129065707001093.
[5] Rowland, David C., Yasser Roudi, May-Britt Moser, and Edvard I. Moser. 2016. “Ten Years of Grid Cells.” Annual Review of Neuroscience 39 (February): 19–40. https://doi.org/10.1146/annurev-neuro-070815-013824.
[6] Stensola, H. et al. The entorhinal grid map is discretized. Nature 492, 72–78 (2012).
[7] Gardner, Richard J., Erik Hermansen, Marius Pachitariu, Yoram Burak, Nils A. Baas, Benjamin A. Dunn, May-Britt Moser, and Edvard I. Moser. 2022. “Toroidal Topology of Population Activity in Grid Cells.” Nature 602 (7895): 123–128.
[8] Stemmler, Martin, Alexander Mathis, and Andreas V M Herz. 2015. “Connecting Multiple Spatial Scales to Decode the Population Activity of Grid Cells.” Science Advances 1 (11): e1500816. https://doi.org/10.1126/science.1500816.
[9] Bush, Daniel, Caswell Barry, Daniel Manson, and Neil Burgess. 2015. “Using Grid Cells for Navigation.” Neuron 87 (3): 507–20. https://doi.org/10.1016/j.neuron.2015.07.006.

0
推荐
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}