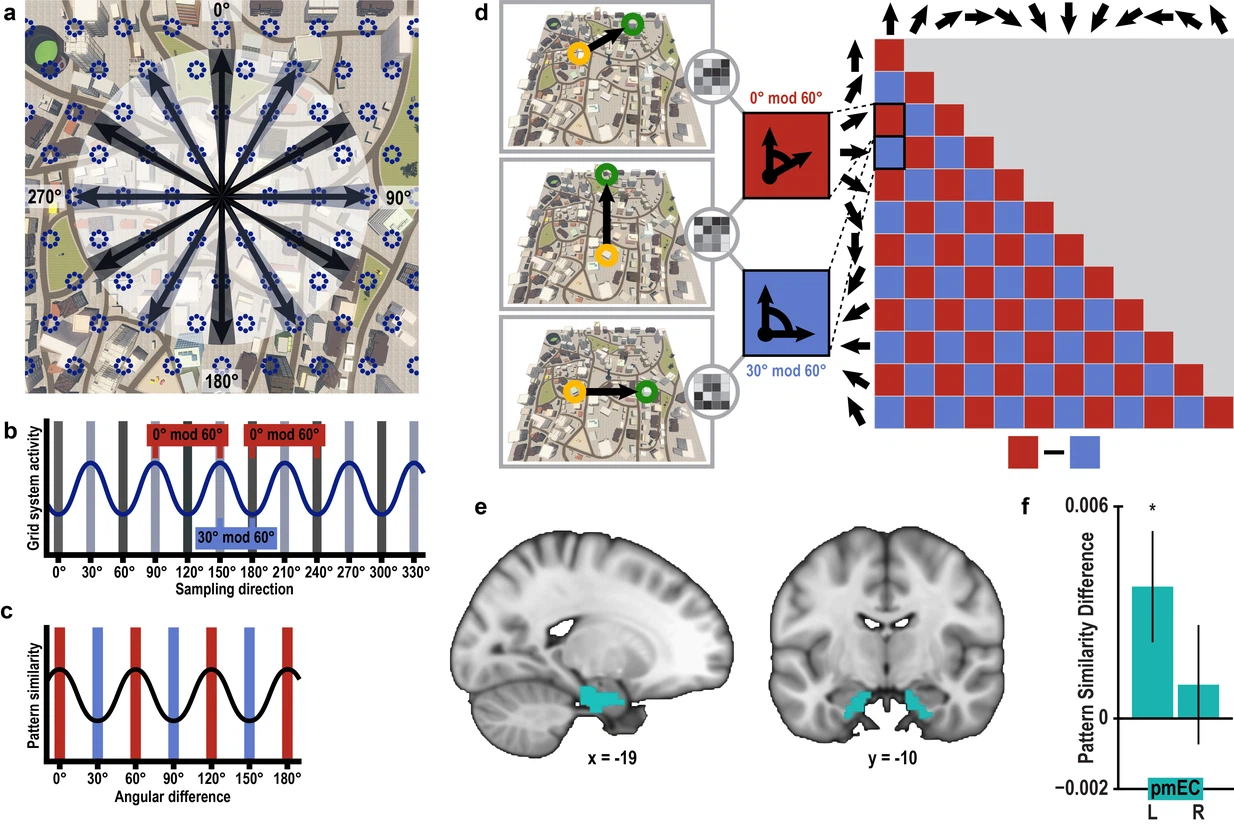
图26.4 志愿者在虚拟城市探索中左侧pmEC呈现六边形网格活动。(a)一个假想的网格细胞及它的备选方向。(b)六边形网格对应60度循环出现的波峰。(c)在fMRI数据中也寻找这样波峰波谷的分布。(d)虚拟城市中从一个点到另一个点的向量,也应遵循相差60度更一致,相差30度更不一致。(e)志愿者fMRI数据中pmEC位置。(f) 左侧pmEC六边形网格活动显著,右侧不显著 (T23 = 0.57, p = 0.58)。来自[18]
参考文献:
[1] Donato, F., Jacobsen, R. I., Moser, M.-B. & Moser, E. I. Stellate cells drive maturation of the entorhinal-hippocampal circuit. Science(80-). 355, eaai8178 (2017).
[2] Gardner, Richard J., Erik Hermansen, Marius Pachitariu, Yoram Burak, Nils A. Baas, Benjamin A. Dunn, May-Britt Moser, and Edvard I. Moser. 2022. “Toroidal Topology of Population Activity in Grid Cells.” Nature 602 (7895): 123–128.
[3] Wang, T. et al. Modularization of grid cells constrained by the pyramidal patch lattice. iScience 24, 102301 (2021).
[4] Guanella, A. & Verschure, P. F. M. J. Prediction of the position of an animal based on populations of grid and place cells: a comparative simulation study. J. Integr. Neurosci. 6, 433–46 (2007). [5] Krupic, Julija, Marius Bauza, Stephen Burton, Caswell Barry, and John O’Keefe. 2015. “Grid Cell Symmetry Is Shaped by Environmental Geometry.” Nature 518 (7538): 232–235. https://doi.org/10.1038/NATURE14153.
[6] Whittington, James C.R., Timothy H. Muller, Shirley Mark, Guifen Chen, Caswell Barry, Neil Burgess, and Timothy E.J. Behrens. 2020. “The Tolman-Eichenbaum Machine: Unifying Space and Relational Memory through Generalization in the Hippocampal Formation.” Cell 183 (5): 1249-1263.e23.
[7] Gu, Y. et al. A map-like micro-organization of grid cells in the medial entorhinal cortex. Cell 175, 736-750.e30 (2018).
[8] Ray, Saikat, Robert Naumann, Andrea Burgalossi, Qiusong Tang, Helene Schmidt, and Michael Brecht. 2014. “Grid-Layout and Theta-Modulation of Layer 2 Pyramidal Neurons in Medial Entorhinal Cortex.” Science (New York, N.Y.) 343 (6173): 891–896. https://doi.org/10.1126/SCIENCE.1243028.
[9] Naumann, Robert K., Saikat Ray, Stefan Prokop, Liora Las, Frank L. Heppner, and Michael Brecht. 2016. “Conserved Size and Periodicity of Pyramidal Patches in Layer 2 of Medial/Caudal Entorhinal Cortex.” Journal of Comparative Neurology 524 (4): 783–806. https://doi.org/10.1002/cne.23865.
[10] Tang, Qiusong, Andrea Burgalossi, Christian Laut Ebbesen, Saikat Ray, Robert Naumann, Helene Schmidt, Dominik Spicher, and Michael Brecht. 2014. “Pyramidal and Stellate Cell Specificity of Grid and Border Representations in Layer 2 of Medial Entorhinal Cortex.” Neuron 84 (6): 1191–97. https://doi.org/10.1016/J.NEURON.2014.11.009.
[11] Beed, P. et al. Analysis of excitatory microcircuitry in the medial entorhinal cortex reveals cell-type-specific differences. Neuron 68, 1059–1066 (2010).
[12] Rueckemann, J. W., Sosa, M., Giocomo, L. M. & Buffalo, E. A. The grid code for ordered experience. Nat. Rev. Neurosci. 22, 637–649 (2021).
[13] Diehl, Geoffrey W, Olivia J Hon, Stefan Leutgeb, and Jill K Leutgeb. 2017. “Grid and Nongrid Cells in Medial Entorhinal Cortex Represent Spatial Location and Environmental Features with Complementary Coding Schemes.” Neuron 94 (1): 83-92.e6. https://doi.org/10.1016/j.neuron.2017.03.004.
[14] Chaudhuri, Rishidev, Berk Gerçek, Biraj Pandey, Adrien Peyrache, and Ila Fiete. 2019. “The Intrinsic Attractor Manifold and Population Dynamics of a Canonical Cognitive Circuit across Waking and Sleep.” Nature Neuroscience 22 (9): 1512–20. https://doi.org/10.1038/s41593-019-0460-x.
[15] Stensola, Hanne, Tor Stensola, Trygve Solstad, Kristian FrØland, May Britt Moser, and Edvard I. Moser. 2012. “The Entorhinal Grid Map Is Discretized.” Nature 492 (7427): 72–78.
[16] Zutshi, Ipshita, Jill K Leutgeb, and Stefan Leutgeb. 2017. “Theta Sequences of Grid Cell Populations Can Provide a Movement-Direction Signal.” Current Opinion in Behavioral Sciences 17 (October): 147–154. https://doi.org/10.1016/j.cobeha.2017.08.012.
[17] Bush, Daniel, and Neil Burgess. 2020. “Advantages and Detection of Phase Coding in the Absence of Rhythmicity.” Hippocampus 30 (7): 745–62. https://doi.org/10.1002/hipo.23199.
[18] Bellmund, J. L., Deuker, L., Navarro Schröder, T. & Doeller, C. F. Grid-cell representations in mental simulation. Elife 5, e17089 (2016).

 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}