从昆虫、鸟类到鲸鱼和人类,歌唱一直是一项社交活动。一般来说,包括人类在内的动物使用较低的音调来彰显首领地位,而使用较高的音调来表示顺从;而不完美则表示有感情,调音太完美了反而不打动人[1]。这可以解释为什么我们有时会觉得人工智能产生的音乐因其完美无缺而显得没有真情实感,实际上更早时候为歌手录制曲目时已经有这样的问题。
音乐作为一种多维输入信息,也是在一开始就被大脑的不同模块处理着。除了音调高低、节奏快慢等基本信息[3],每一小段音乐都可能让我们想起一些场景,或激活一些情绪。这种不同于当前现实的时空情境,可能也涉及一些我们在睡眠章节介绍的脑电波。接受过11个电影片段训练的小鼠,再听到其中的声音,初级视觉皮层(V1)也会活跃,这并非来自听觉皮层的直接输入,而是跟小鼠的细微动作有关[4]。
大鼠还可以练习“演奏”。实验中大鼠被训练按下操纵杆一定时间后释放,以在一个连续的频率轴上播放出特定高频段的声音,这项任务与前文在空间探索章节讲到的类似,也涉及包括“位置细胞”在内的海马体神经元和包括“网格细胞”在内的内侧内嗅皮层(MEC)神经元[5]。这与笔者认为这是一套在大脑皮层普遍投射的搜索、生成、迭代算法相一致(图32.2),我们在因果推断的章节会继续讲。
而成年人可能意识不到,准确地发声是一项全身性的协调运动。小鼠脑干后疑核(nucleus, retroambiguus,其中的约160个表达神经降压素neurotensin的神经元)通过协调下巴、舌头、声带、呼吸等来控制发声的气流,调节音量(图35.1)[6]。这些神经元接受电刺激后对口咽和呼气的肌肉输出的信号是高保真、短延迟的。后疑核中的神经元控制声带,还可以发出小鼠标志性的超声(2万赫兹以上),并与呼吸的相位协调、产生音节[6,7]。
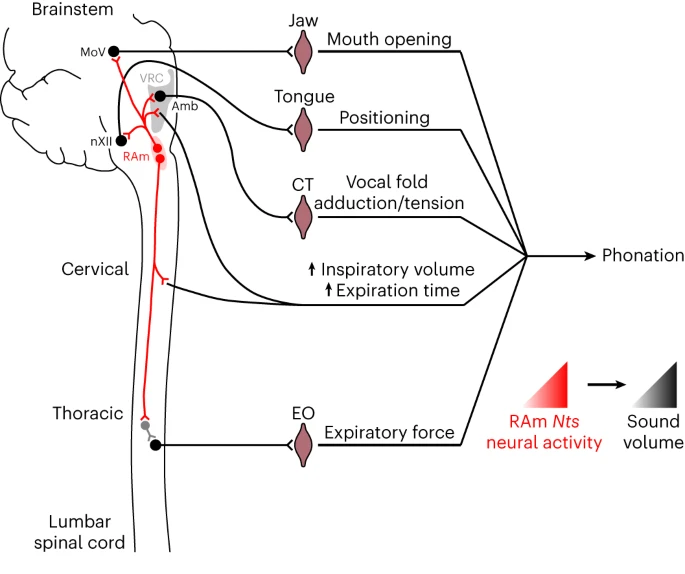
图35.1 小鼠脑干后疑核中部分神经元调节发声的音量和频率。CT,cricothroid,环甲肌,控制声带。EO,external oblique,腹外斜肌,腹式呼吸。来自[6]
人类过了婴儿哺乳期后,喉部逐渐下移,不再像黑猩猩那样发出高频声音[1]。这被认为为复杂语言提供了更多可能性。当前几大语种的元音可能主要反映了嘴张的大小(英语如图35.2c,由Mid+low和High表示)。而在干燥环境中可能信息量更大的辅音,则更为多样,涉及舌头、嘴唇、颚等的不同位置(图35.2)。这其实也是在有限的几个选项中选则一个,因此预计也涉及“网格细胞”。一项对前额叶的研究发现了不同发音对应的神经元细胞(图35.2)[8]
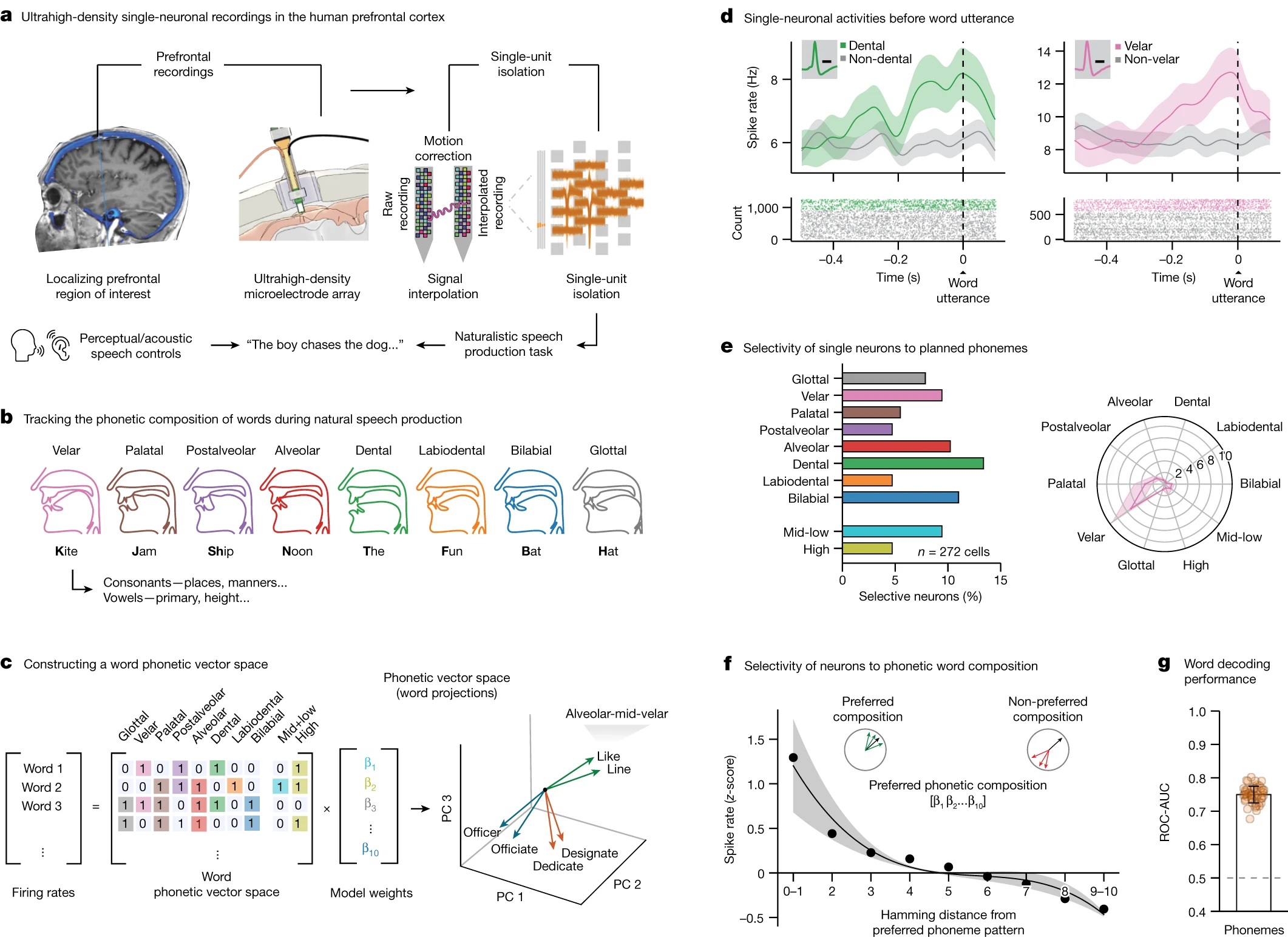
图35.2 接受脑深度刺激电极植入的英语母语的右撇子人的大脑前额叶神经元与发音。(a)实验示意图。(b)发音对应的嘴型,辅音分为8类。(c)每个单词由8个辅音音位和2个元音音位形成向量。(d)前额叶中单个神经元在发音时的放电情况。(e)神经元特异性的分布。左侧为在272个神经元中占比,右侧为单个神经元在10个音位盘上的调谐曲线。(f)标准分数平均的神经元放电率与发音偏好。汉明距离(Hamming distance)每差一个音位记为1,离该神经元对应的单词距离越远,神经元越不放电。(g)根据神经元活动预测单词。来自[8]
在雄性阿尔斯顿歌唱鼠(Alston’s singing mouse)其口面部运动皮层(Orofacial Motor Cortex, OMC)的研究中,发现OMC区域短时间尺度(约100毫秒)的活动可能反映的是感官反馈,而长时间尺度(10秒)的放电活动会有“拉伸”或“压缩”[9]。这可能是一种相对长程的范式的运用。
发声的社交性可以影响个体的行为表现。当引入关于音高这一反馈时,大提琴手会仔细聆听,但是不会被反馈影响演奏,而女高音歌手则会用她们脑中与音乐相关的区域来补偿这些扰动,即使她们唱得本来没问题[10]。群居的野生长臂猿也会协调其发声,雌性协调适应雄性以保持节奏促成合唱[11]。当一起唱歌时,唱歌的鸟儿也会相互协调,跟独自唱的时候不一样,还可以看到较为长程的结构(图35.3)。
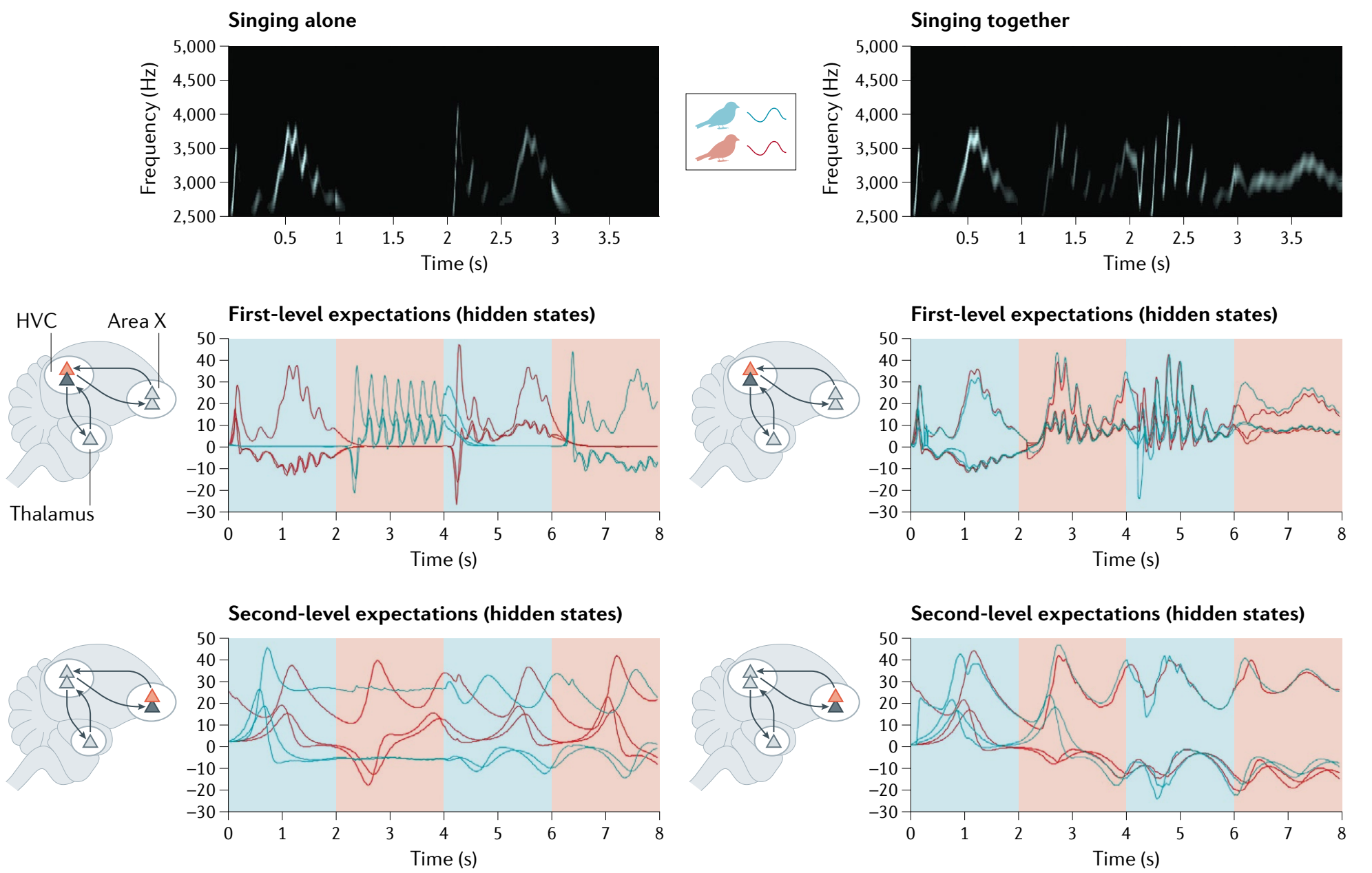
图35.3 利用鸟鸣进行通信。两只具有相同生成模型但初始条件不同的鸟(A)轮流唱歌2秒,然后倾听回应(B)。阴影区域表示当前正在唱歌的鸟:红色表示第一只鸟,蓝色表示第二只鸟。唱歌时,感官预测误差被抑制,以便通过行动实现预测。相反,倾听时,感官预测误差通过以更高精度采样,从而被关注。上面的面板显示了第一只鸟听到的声谱图(下面的面板中的红线;请注意,上下面板的时间尺度不同)。在(A)中,两只鸟无法彼此听到,而在(B)中,它们可以彼此听到。第一只鸟(红色)的后验期望以时间为函数显示在上方——第二只鸟(蓝色)的相应期望以蓝色显示。在左侧子图中,由于这只鸟只能听到自己,声谱图反映了基于高级发声肌(HVC;中间子图中的前运动区)和区域X(更高级别的区域,下部子图)的期望的下降性本体预测。中间和下部面板中分别报告了关于生成鸟鸣的基础原因(即振幅和频率的波动)的HVC和区域X的期望。请注意,当鸟儿倾听时,它们在第一级上的期望会降至零,因为它们听不到任何东西。然而,区域X较慢的动力学可以在每次倾听期结束后再次生成歌曲。在右侧子图中,两只鸟可以听到彼此。在这种情况下,在感官和超感官层次上都显示出同步的期望。请注意,声谱图在连续的2秒时间段内生成——交替由第一只鸟和第二只鸟生成。精度的关键作用再次显现;在这里,选择性地关注由鸟儿产生的感官流——以一种协调的方式,使它们能够轮流唱歌和交流。来自[3]。
在基本了解了多要素的发声调控,和在此基础上相对复杂的组合后,我们在下一节会进一步探讨口头语言和书面语言的神经基础。
参考文献:
[1] Colapinto, John. 2021. This Is the Voice. New York: Simon & Schuster.
[2] Segado, Melanie, Robert J Zatorre, and Virginia B Penhune. 2021. “Effector-Independent Brain Network for Auditory-Motor Integration: FMRI Evidence from Singing and Cello Playing.” NeuroImage 237: 118128. https://doi.org/10.1016/j.neuroimage.2021.118128.
[3] Vuust, Peter, Ole A. Heggli, Karl J. Friston, and Morten L. Kringelbach. 2022. “Music in the Brain.” Nature Reviews Neuroscience 23 (5): 287–305. https://doi.org/10.1038/s41583-022-00578-5.
[4] Bimbard, C., Sit, T.P.H., Lebedeva, A. et al. Behavioral origin of sound-evoked activity in mouse visual cortex. Nat Neurosci 26, 251–258 (2023). https://doi.org/10.1038/s41593-022-01227-x
[5] Aronov, D., Nevers, R. & Tank, D. Mapping of a non-spatial dimension by the hippocampal–entorhinal circuit. Nature 543, 719–722 (2017). https://doi.org/10.1038/nature21692
[6] Veerakumar, A., Head, J.P. & Krasnow, M.A. A brainstem circuit for phonation and volume control in mice. Nat Neurosci 26, 2122–2130 (2023). https://doi.org/10.1038/s41593-023-01478-2
[7] Park J, Choi S, Takatoh J, Zhao S, Harrahill A, Han BX, Wang F. Brainstem control of vocalization and its coordination with respiration. Science 383, eadi8081 (2024)
[8] Khanna, A.R., Muñoz, W., Kim, Y.J. et al. Single-neuronal elements of speech production in humans. Nature 626, 603–610 (2024). https://doi.org/10.1038/s41586-023-06982-w
[9] Banerjee, A., Chen, F., Druckmann, S. et al. Temporal scaling of motor cortical dynamics reveals hierarchical control of vocal production. Nat Neurosci 27, 527–535 (2024). https://doi.org/10.1038/s41593-023-01556-5
[10] Segado, Melanie, Robert J Zatorre, and Virginia B Penhune. 2021. “Effector-Independent Brain Network for Auditory-Motor Integration: FMRI Evidence from Singing and Cello Playing.” NeuroImage 237: 118128.
[11] Ma, H., Wang, Z., Han, P., Fan, P., Chapman, C. A., Garber, P. A., & Fan, P. (2024). Small apes adjust rhythms to facilitate song coordination. Current Biology, 34(5), 935-945.e3. https://doi.org/10.1016/j.cub.2023.12.071

0
推荐
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}