阅读:0
听报道
在细说记忆之前,需要先介绍一下主要感官,它们决定了记忆和检索模块的输入信息。不同于逐层“汇报”,成年动物的感官在接受外界刺激的时候,接受感官刺激的细胞自身就在做信号处理。感官会对输入数据进行稀疏化,而单一功能是通过相对较少的神经元来实现的。
从海洋动物、昆虫到人类,我们都受到了我们感官的时空分辨率的限制和保护,而时空分辨率是我们记忆和反应的基础。一个生物体只需获得了它需要做什么的必要信息(例如,细菌的趋化性),更多的信息并不一定会更好。这是所有感官都需要遵守的一般原则。
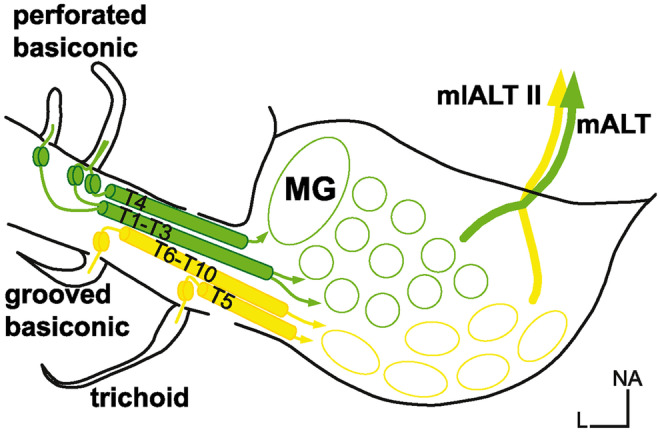
图8.1 昆虫体内嗅觉信号的分离。(A)蟑螂大脑从外周到高级脑中枢的嗅觉通路示意图。孔状的基底感觉器(绿色)的嗅觉感觉神经元(OSNs)穿过触角束(antennal tracts)T1-T4,终止于前腹侧的小球(绿色)。在这些小球中,带有树突的投射神经元(PNs)通过触角与脑之间的mALT束(mediolateral Antenno-cerebral tracts)投射到前脑。毛状和槽状基底感受器(黄色)中的嗅觉感觉神经元分别穿过触角束T5和T6-10,终止于后背侧的小球(黄色)。在这些小球中带有树突的投射神经元通过mlALT II束投射到前大脑,来自[1]。
嗅觉是最古老的一种感官,在无脊椎动物(如蠕虫、昆虫)和脊椎动物中都起着关键作用(从鱼类到人类)(图8.2)。在蟑螂中,包括信息素在内的嗅觉信号,在进入大脑之前,在触角中被局部分离和处理 (图8.1)。从进化上来看,嗅觉可以追溯到细菌的化学感应(例如感知周围环境中食物或有害物的浓度),这种感应能力被所有生物继承,被用以探索其所在环境。对于动物来说,嗅觉在自动跟踪任务的应用,可能会比视觉更经济。在日常以果蝇为主的广东地区,几块臭豆腐就可以让多只苍蝇从不知道多少米之外赶来。鱼类的嗅觉可能也有我们很难想象的远距离推断功能。大脑(前脑)紧跟在嗅球之后(图8.2)绝非偶然。
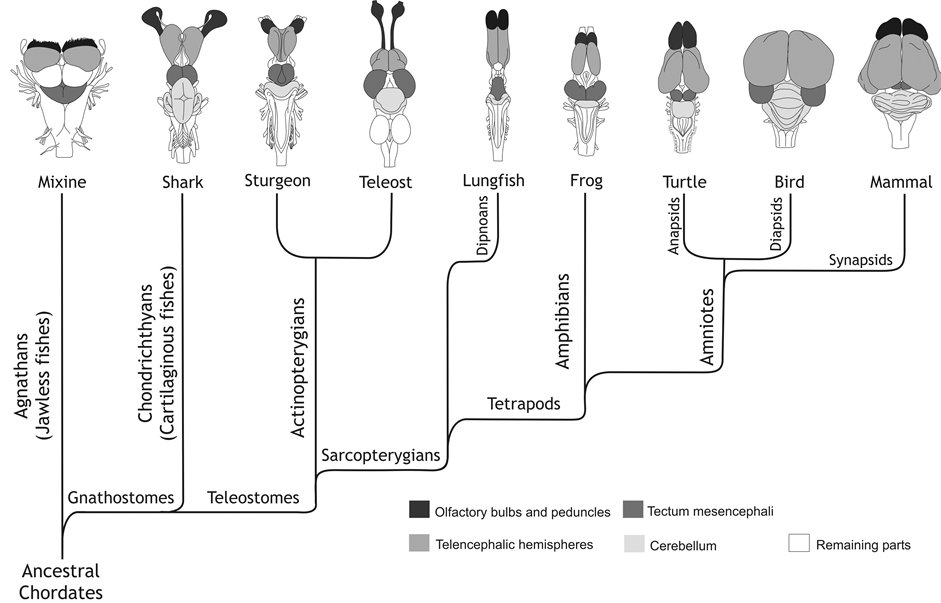
图8.2 脊椎动物嗅球(黑色)、前脑、中脑、后脑形态的顶视图。来自[2],本书开篇也有出现。
一般来说,一次学习一件事在动物中是一种普遍模式。这不同于常用的试图在两个(或更多的)组之间进行分类的有监督学习算法。果蝇体内约2000个 凯尼恩(Kenyon)细胞,每种气味大约激活其中5%的细胞(图8.3)。
具体来看: 50个气味分子对应了果蝇的50个投射神经元,每个投射神经元的连接方式是“随机但重叠”的。图7.3 B中的每个分子,可被看成对应一个长度50的向量,随机性意味着不同气味会激活不同的PN组合,形成一种“局部哈希”编码。来自投射神经元的信息多对一地到达凯尼恩细胞后,赢者通吃的机制保证了特定凯尼恩细胞对APL神经元的激活,当多个PN同时激活时,只有最强那个凯尼恩细胞被激活,其余被抑制APL细胞,通过反馈决定了对特定气味的爱憎(attraction-aversion)决策 [3]。
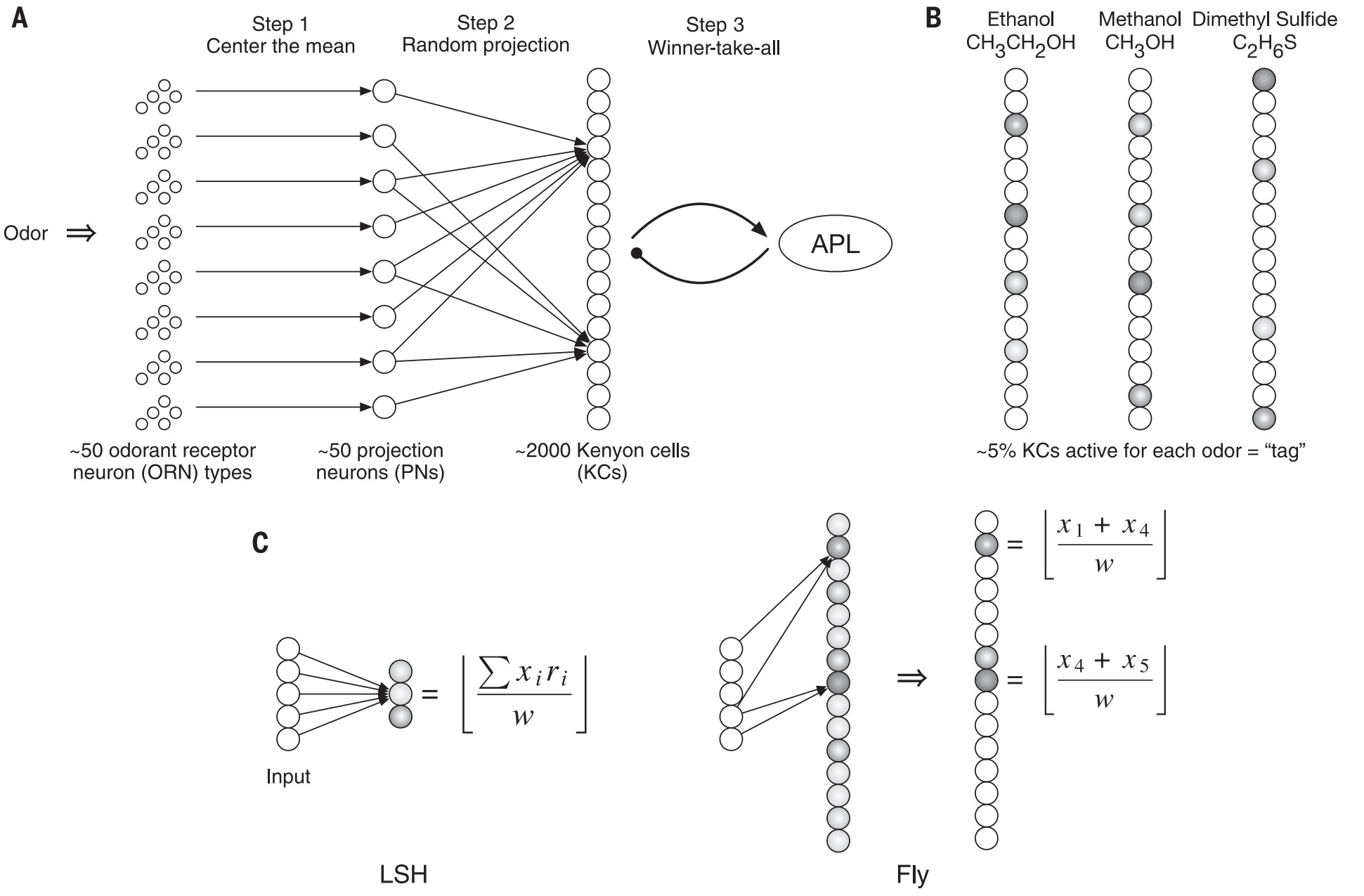
图8.3 苍蝇嗅觉回路和局部敏感哈希(LSH)对比。(A)苍蝇嗅觉回路示意图。(B)气味反应的示意图。相似的气味对(如甲醇和乙醇)被分配了更相似的标签。(C)传统的局部敏感哈希(LSH)算法与苍蝇嗅觉算法的区别。来自[3]。
在果蝇中,中间神经元具有增益控制功能。在缺乏足够多可用的神经元时,果蝇的嗅觉不同于前述的局部敏感哈希,这些抑制性神经元之间的相互交互,可减少信号失真并提高刺激辨识能力。该案例可视为使用多个串联的反馈回路,来适应性地调节网络输出的一个特例,在人工智能中也可借鉴[4]。
接下来介绍几个基于嗅觉机制改进神经网络网络的例子:果蝇嗅觉网络的结构(图8.3)使终身持续学习成为可能,比现有的使用深度神经元网络(如梯度情景记忆(GEM)、弹性权重巩固(EWC)和脑启发重放)的持续学习方法会更少的遇到灾难性遗忘[5]。要让人工神经网络能够具有持续学习能力,可借鉴嗅觉处理中保护神经元已有信息,避免灾难性重写的机制。
除了持续学习之外,果蝇嗅觉网络已被证明可以用于自然语言处理,与主流算法,如(GloVe)相比,具有更快的训练时间和更少的存储消耗[6]。
另一项基于曼杜卡六角蛾(Manduca sexta Moth)嗅觉学习的计算模型[7],指出构建从非常少的样本快速学习的神经网络,可借鉴的机制包括高噪声层,作为噪声过滤器的稀疏层,以及基于章鱼胺(octopamine )刺激,稀疏层和基于赫布法则的网络训练方法。
基于昆虫嗅觉的特征提取器MothNet[8],借鉴了昆虫嗅觉中的竞争性抑制层,高维稀疏表征和基于赫布法则的权重更新,构建特征预处理器。在视觉数据集MINST上,MothNet 特征生成器的性能也大大优于其他特征预处理·方法,如 PCA、 PLS 和神经网络。取决于基线分类算法的准确度,可提升6%-33%的算法性能。
嗅觉从进化上来看,首要是为了寻找配偶,也可用找出食物,这都需要利用化合物之间的联系,尤其是当多种化合物是来自同一来源时。为了节省存储空间,共同出现的分子倾向于激活同一个神经元,而如果两种分子很少共同出现,神经系统就应该让其各自对应的神经元放在距离相对更远的地方,如此可创建了一个三维双曲空间(一个呈负曲率的球体,类似马鞍)地图[9]。这种几何上的匹配可以避免将气味映射到感知时产生的扭曲,与视觉系统处理图像时,根据像素相似度在双曲空间创建的地图类似。
气味浓度的梯度变化是包括人类在内的动物进行空间导航所需的关键信息。两个鼻孔的气味浓度差异(对于人类,该值为3.5厘米)可以进行比较。在人类中,来自两个鼻孔的信号也分别到达嗅觉皮层,来自于同侧的信号比来自不同侧的信号快大约480毫秒[10]。左右之间的差距可用于定位与导航,对应的证据是如果一个鼻孔被阻塞,盲鼹鼠最终会在绕了更多的路之后找到气味来源,搜寻方向通常更多地朝向鼻孔畅通的一侧[11](图8.4)。
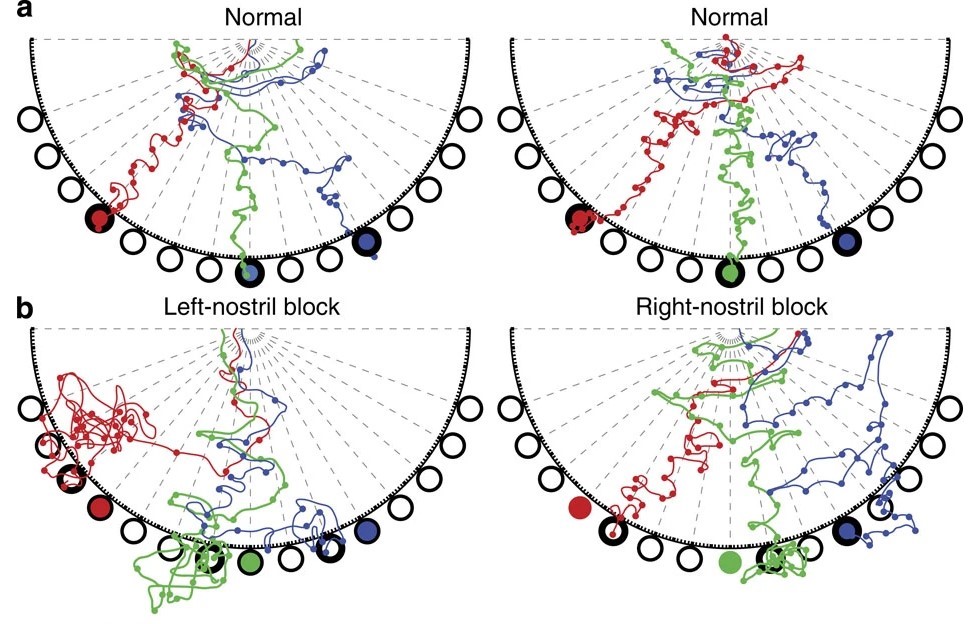
图8.4 在正常和鼻孔阻塞条件下的盲鼹鼠搜索路径。(A)两只盲鼹鼠的正常搜索行为。搜索的颜色与每次试验的目标井的颜色相匹配。圆圈表示嗅探物。每个实验中第一个用鼻子倾斜探索的井用粗体轮廓表示。(B)(A.中显示的左、右鼻孔块的搜索行为示例左鼻孔阻塞偏向右搜索,而右鼻孔阻塞偏向左搜索。彩色的井表示目标(食物)位置;粗体的轮廓表示第一个探索的探索。痕迹被说明,直到目标井被探索,鼹鼠会持续搜索,并在几乎每次试验中都可找到食物。来自[11]。
图8.4中嗅探后的每一个回合都就像梯度学习的一步(在人工智能中),更接近气味源时,嗅探的步长更短(图8.4)。深度学习中Adagrade算法中,控制权重变化大小的参数学习率(learning rate),只是随着梯度的偏导数降低而降低。而盲鼹鼠在这里展示的,更可能是一个经过对气味浓度比较的趋近过程,高浓度意味着已经相对接近目标,不应作大幅调整。两者在机制上有所不同。
在复杂的空气动力学条件下,气味浓度的变化频率可以达到1000赫兹。连续嗅探(例如,每秒10次,10赫兹)并没有跟得上自然界气味浓度的时间变化速度。网上能看到的一些累趴了的工作犬就往往是因为连续嗅探。有证据说明小鼠能够区分高达40赫兹[12]的气味频率。后面章节讲到的脑电波的相位信息可能被用来区分这样的高频变化。脑电波的相位信息,相当于为高频气味变化增加了一层可调的正弦函数,以实现高于嗅探频率的高分辨率。
嗅觉回路可以检测两种分子之间的正相关性和负相关性,这是迈向检测因果关系的第一步,检测相关性也可用于分离气味的来源。
参考文献:
[1] Fuscà, D. & Kloppenburg, P. Odor processing in the cockroach antennal lobe-the network components. Cell Tissue Res. 383, 59–73 (2021).
[2] Broglio, C. et al. Hippocampal Pallium and map-like memories through vertebrate evolution. J. Behav. Brain Sci. 05, 109–120 (2015).
[3] Dasgupta, S., Stevens, C. F. & Navlakha, S. A neural algorithm for a fundamental computing problem. Science (80-.). 358, 793–796 (2017).
[3] Dasgupta, S., Stevens, C. F. & Navlakha, S. A neural algorithm for a fundamental computing problem. Science (80-.). 358, 793–796 (2017).
[4] Barth-Maron, A., D’Alessandro, I., & Wilson, R. I. (2023). Interactions between specialized gain control mechanisms in olfactory processing. Current Biology : CB, 33(23), 5109-5120.e7.
[5] Wang, L., Zhang, X., Li, Q., Zhang, M., Su, H., Zhu, J., & Zhong, Y. (2023). Incorporating neuro-inspired adaptability for continual learning in artificial intelligence. Nature Machine Intelligence, 5(12), 1356–1368.
[6] Bao, Pinglei, Liang She, Mason McGill, and Doris Y Tsao. 2020. “A Map of Object Space in Primate Inferotemporal Cortex.” Nature 583 (7814): 103–108.
[7] Delahunt, C. B., Riffell, J. A., & Nathan, K. J. (2018). Biological Mechanisms for Learning: A Computational Model of Olfactory Learning in the Manduca sexta Moth, with Applications to Neural Nets. ArXiv.org. https://arxiv.org/abs/1802.02678
[8] Delahunt, C. B., & Nathan, K. J. (2018). Insect cyborgs: Bio-mimetic feature generators improve machine learning accuracy on limited data. ArXiv.org. https://arxiv.org/abs/1808.08124
[9] Zhou, Y., Smith, B. H., & Sharpee, T. O. (2018). Hyperbolic geometry of the olfactory space. Science Advances, 4(8). https://doi.org/10.1126/sciadv.aaq1458
[10] Gülce Nazlı Dikeçligil, Yang, A. I., Sanghani, N., Lucas, T., H. Isaac Chen, Davis, K. A., & Gottfried, J. A. (2023). Odor representations from the two nostrils are temporally segregated in human piriform cortex. CB/Current Biology, 33(24), 5275-5287.e5.
[11] KC, Catania. 2013. “Stereo and Serial Sniffing Guide Navigation to an Odour Source in a Mammal.” Nature Communications 4. https://doi.org/10.1038/NCOMMS2444
[12] Ackels, Tobias, Andrew Erskine, Debanjan Dasgupta, Alina Cristina Marin, Tom P A Warner, Sina Tootoonian, Izumi Fukunaga, Julia J. Harris, and Andreas T. Schaefer. 2021. “Fast Odour Dynamics Are Encoded in the Olfactory System and Guide Behaviour.” Nature 593 (7860): 558–63.
话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}